



在论述人类独特思维的演化起源上,我们首先将关注重点放在人类与其他现存哺乳类动物分化之前具有的最后共同祖先上,最好的代表就是人类的近亲——非人类人猿(现在类人猿的祖先),包括黑猩猩、倭黑猩猩、大猩猩、猩猩。其中,尤其是大约600万年前才与人类走上不同演化道路的黑猩猩和倭黑猩猩,更是我们着重要关注的。上述四种类人猿在认知能力上是相似的,人类却有所不同,我们假设这是由于现存类人猿保持了我们共同祖先的认知技能,人类分化之后却演化出了一些新的东西。
我们对人类与现存类人猿共同祖先认知技能的了解来源于类人猿的实验研究,而这些研究直接促进了我们前面提到的个体意图性(涉及认知模型和工具性推理参与的自我调节及某些类型的自我监控)理论框架的搭建。由于人类与其他类人猿的分化发生于近期(以物种演化的时间进程标定,可从具有的基本相同的躯体结构、感觉器官、情绪及大脑结构上体现出来),在没有直接反对证据的情况下我们假定人类与类人猿在演化上具有连续性(de Waal,1999),也就是说,当类人猿在精确操控的实验中表现出与人类相同的行为时,我们假定背后涉及的认知过程具有连续性。当然,在后面的章节中我们将会提到不同假设条件下思维的解释问题。
类人猿的认知和思维过程可以被分为两类:用于理解充满物理性因果联系的物理世界;用于理解充斥着机体之间因果/意图关系的社交世界。灵长类动物对物理世界的认知主要从觅食情境中演化而来(Tomasello & Call,1997),所以主要服务于这个目的(Millikan,1987)。为了保证每天充足的食物供给,它们演化出了近期目标、表征、推理等内容,服务于:
(1)寻找食物(需要空间定向和目标追踪技能);
(2)识别并对食物进行归类(需要特征识别及归类技能);
(3)确定食物的量(需要量化技能);
(4)获取或提取食物(需要理解事物间因果联系的技能)。
在这些用于理解物理世界的最基本技能的掌握上,所有非人灵长类动物之间并无太大区别(Tomasello & Call,1997;Schmitt et al.,2012)。
与其他灵长类动物相比,类人猿尤为擅长的是对工具的使用,在这一点上我们甚至可以认为它们不仅理解了事物之间的因果联系,甚至可以对这种联系进行操控,相比而言,其他灵长类动物最多在某一特定情境下才会使用工具(Fragaszy et al.,2004)。前面提到的四种类人猿都可以很灵活地使用工具。例如,在同一任务中连续使用两种工具、把两个工具连接在一起以便够取食物,等等(Herrmann et al.,2008)。一般认为,工具使用涉及使用者对所使用的工具可能对目标物体或事件产生的影响进行的评估(Piaget,1952),所以类人猿在成功使用新异工具时表现出的灵活性和敏捷似乎表明,他们具有一个或更多能够指引其在新异工具使用时进行因果关系推论的一般认知模型。
类人猿借助工具对因果联系进行操控的技能与它们的认知表征和推理过程之间可能存在着有趣的联系。例如,马林·曼瑞奇等人(Marín Manrique,Cross & Call,2010)在实验中给黑猩猩呈现一个从未遇到过的食物获取问题,该问题的解决需要一个具有特定属性(如需要具有一定的硬度和强度)的工具才能完成。然而,在实验中,可用工具却被放在另一间不同的房间。为了解决这个问题,黑猩猩首先必须理解问题情境中包含的因果结构,之后形成对这种结构的认知表征并保存下来,以便到另一间房间寻找工具时使用。研究结果发现,往往从第一次尝试开始,很多黑猩猩就是这样做的,这也意味着它们可以把该新问题同化到已有包含因果结构的认知模型当中,并带着该认知模型进行后续行为(到隔壁房间),之后借助该认知模型在头脑中模拟工具的使用方法及可能带来的结果。在马尔卡希和卡尔(Mulcahy & Call,2006)的研究中,倭黑猩猩甚至可以为未来储存工具,这大概是因为它们认为未来遇到相关情境时可以进一步使用。
这里提到的模拟和推理是具有逻辑结构的。然而,这种逻辑结构并非指形式逻辑结构,而是基于因果推理产生的结构。这里的观点是:因果推理遵循基本的“如果-那么”逻辑,因此,可以从中推出“必然”结论,即如果A发生,那么B发生(因为A导致B)。伯慕德兹(Bermudez,2003)称这类推理为“原型条件性”(protoconditional)推理,因为这里涉及的核心不是形式上的,而是因果性的。在曼瑞奇等人(Manrique,Cross & Call,2010)的实验中,当类人猿通过不同工具模拟时,它会推断“如果使用具有A属性的工具,B会发生”,这样个体就逐渐获得假言推理(modus ponens)原型,之后在使用具有A属性工具时就会期望B作为结果发生(如果A发生,那么B发生;A发生了所以B将会发生)。这是一个基本的从前提/原因到结论/后果的正向推理过程。
在近期进行的另一类研究中,研究者亦发现了从结果到原因的逆向推理。在卡尔(Call,2004)的实验中,首先向黑猩猩展示一块食物,之后把食物藏进两个杯子中的其中一个里面(黑猩猩不知食物被具体藏在了哪个杯子里面),然后,依据实验条件,实验者摇晃其中的一个杯子。这有助于黑猩猩成功找到如下食物的背景信息:
(1)食物被藏在两个杯子中的其中一个里面(通过之前的训练习得);
(2)摇晃放有食物的杯子会发出声响,摇晃没有放置食物的杯子则不会发出声响(实验中包含的因果性知识)。
在图2-2[3]中,我们以符号化呈现的方式描述了类人猿理解该问题情境的可能方式。
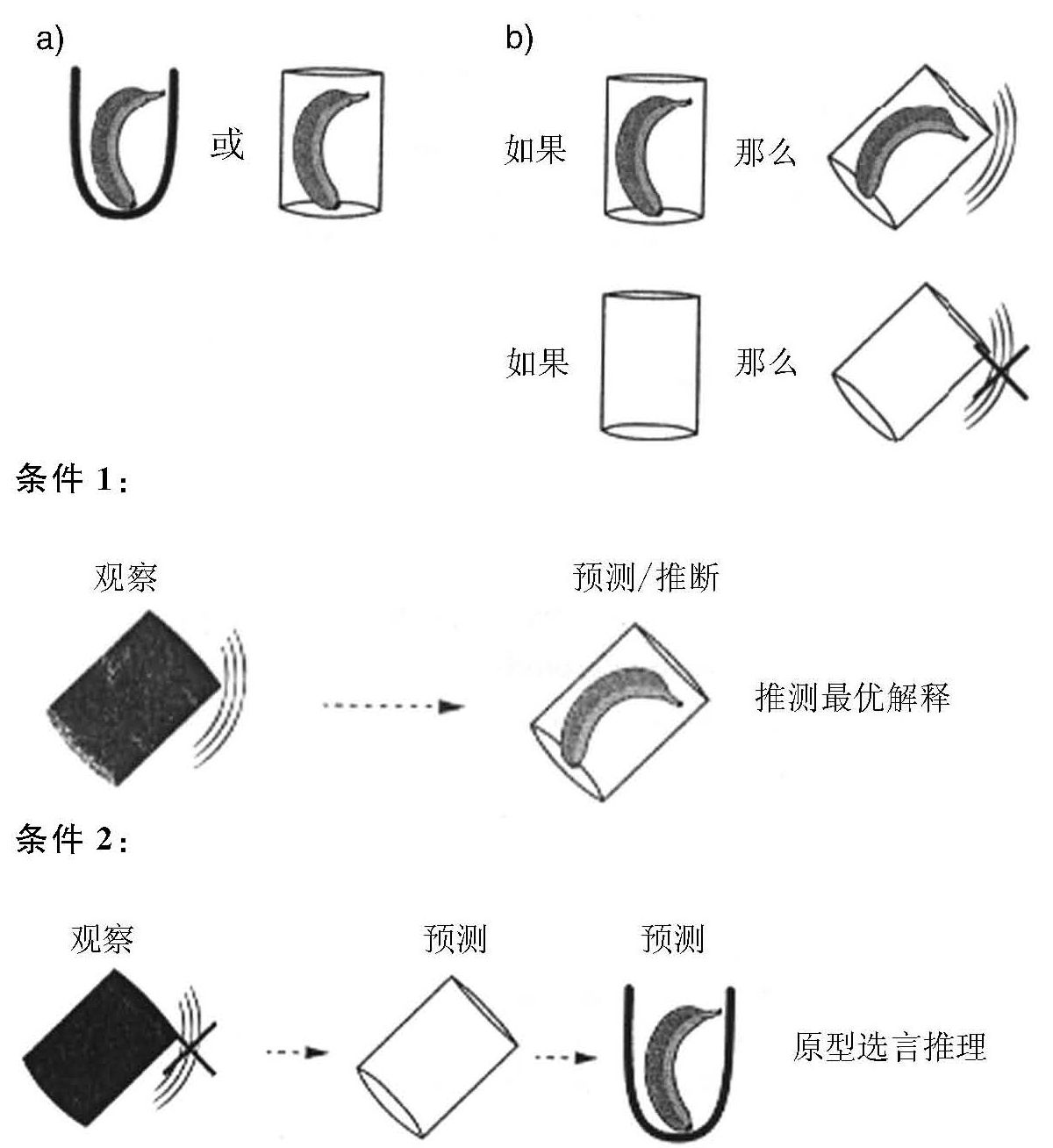
图2-2 寻找隐藏食物时类人猿的推理(Call,2004)
在条件1中,实验者摇晃装有食物的杯子。这种情况下黑猩猩会听到杯子发出声响,并对发出声响的原因(尤其是食物撞击杯子内壁这一原因)进行逆向推理,这是一种不明推论(不符合逻辑,但却是最佳解释推论),遵循的思路是:
(1)摇晃杯子时产生了声响;
(2)如果食物在杯子里,那么摇晃杯子的时候会发出声响;
(3)因此,食物在杯子里。
在条件2中,实验者摇晃没有放置食物的杯子,这种情况下黑猩猩听不到声响,并逆向推理为什么会这样(杯子里没有食物),这是一种原型否定式推理(proto-modus tollens),遵循:
(1)摇晃杯子,没有发出声响;
(2)如果食物在杯子里,那么摇晃杯子的时候会发出声响;
(3)因此,食物一定不在杯子里(杯子是空的)。
值得注意的是,除了进行上述推理,黑猩猩还会进行一个额外推理,在这个例子中,它会把自己对声响产生因果关系的理解与食物被放置在两个杯子中的其中一个这条信息联合起来,以在没有摇晃另一只(other)杯子的情况下确定杯子里是否有食物。这样,黑猩猩的这种推论就带有析取三段论中的排除性推理成分。
“否定”是很复杂的认知操作,因此“类人猿逻辑推理时涉及‘否定’过程”这类说法很容易就会受到攻击。伯慕德兹(Bermudez,2003)关于形式否定出现之前的一些逻辑形式先导的论述使得上述说法相对变得更容易被接受,他把原型否定(protonegation)当作包含互斥构念的一个连续体,如“在-不在”“嘈杂-安静”“成功-失败”“可用-不可用”。按照此种说法,如果我们假设类人猿可以理解这些互斥构念(例如,如果某些东西缺失,它们不可能同时在场;发出噪声的同时不可能保持安静),那么否定的发生就成为可能。实际上,我们这里谈及的“否定”都是从伯慕德兹对原型否定的定义这一层次来说的。
条件性操作(如果-那么)和否定操作组成了人类逻辑推理的基本成分,同时类人猿有能力通过:
(1)已有包含因果结构的认知模型同化潜在问题情境中的核心成分;
(2)包含原型条件性及原型否定成分的正向或逆向推理模型,模拟或推断过去已经或将来将要发生什么,来解决复杂而且以前没有遇到过的物理性问题。
基于上述考量,我们认为,既然研究中类人猿可以使用包含因果规则的认知模型,并可以在自我监控的参与下按照多种不同原逻辑范式(protological paradigm)进行模拟和推理,因此,它们在这些实验任务中的表现应该属于思维范畴。
灵长类关于社交世界的认知主要从群体中为了争夺食物、配偶以及其他有价值的资源而产生的竞争性环境中演化而来(Tomasello & Call,1997),因此,它主要服务于竞争性社会互动。为了在与其他群体成员的竞争中占据优势,灵长类个体演化出了近期目标、表征和推理,以服务于:
(1)识别同群体中的其他个体,并同他们形成某种关系(支配或依从);
(2)识别第三方个体彼此之间的关系,如双亲、上司、朋友之间的关系,并把这些关系纳入自己的认知框架。
这些能力使个体在复杂的社会环境中能够更好地预测其他个体的行为(Kummer,1972)。尽管社会结构和社会互动在不同物种之间存在很大的区别,但在社会认知的基础技能方面,所有灵长类动物都具有很大程度的形似性(Tomasello & Call,1997;Mitani et al.,2012)。
除了具有通过观察社会互动识别社会关系的能力外,类人猿还能把其他个体当作一个具有追求目标和知觉注意情境的主体来看待,进而知道个体的目标和知觉,进一步决定其行为。这也意味着类人猿不仅本身具有意图,同时还可以将其他个体理解为具有意图的个体(Call & Tomasello,2008)。
下面我们一起来看一下相关实验。黑尔等人(Hare et al.,2000)在实验中创设了一个竞争性实验条件,实验中两只黑猩猩都想获取食物,但它们的地位不同,一只处于支配地位,另一只则处于从属地位,同时实验中包含两处食物,一处被放置在开放空间,两只黑猩猩都能看到;另一处则被放到障碍物后面,只有处于从属地位的黑猩猩能够看到。在这种情境下,处于从属地位的黑猩猩知道处于支配地位的黑猩猩能够看到开放空间里的食物,并且有机会时它会尽快获取该食物,然而它却看不到另一块食物(只会看到障碍物),所以也不会想要获取它。实验结果发现,当处于从属地位黑猩猩的笼门被打开后(比处于支配地位黑猩猩笼门打开的时间稍微提前一些),它会选择获取那块被挡在障碍物后面的食物,或许它知道处于支配地位的黑猩猩能够看到什么和不能看到什么。在该实验的一个重要变式中,如果处于支配地位的黑猩猩看到了食物放置的整个过程,虽然之后从它的视角不能看到藏在障碍物后面的食物,但当笼门打开后,处于从属地位的黑猩猩们不会选择去获取那块处于支配地位的黑猩猩“看不到”的食物,它们知道它“知道”食物被放置的位置(Hare et al.,2001;Kaminski et al.,2008)。在另一个变式中,实验者设计了连续进行多次的轮流觅食游戏,黑猩猩们从游戏中能够习得,如果对方首先选择,对方将会选择一个在桌上倾斜放置的木板(似乎下面有些东西),而非平躺放置的木板(下面不会压着其他东西),它们知道在这种情境下对手会做何种推理(Schmelz et al.,2011)。这样看来,黑猩猩不仅知道其他个体看到什么、知道什么,也知道它们如何对事物进行推测。
除了有能力理解其他个体会经验些什么,以及这种经验如何影响它们的行为,类人猿有时甚至试图操纵其他个体的经验。在一系列研究中,研究者创设了黑猩猩与人类个体“争夺”两块食物的问题情境(Hare et al.,2006;Melis et al.,2006a)。在一些条件下,如果黑猩猩接近任何一块食物,实验中的人类个体都可以看到,这时黑猩猩没有表现出接近任何一块食物的行为倾向;在另一些条件下,其中的一块食物被障碍物遮挡,致使黑猩猩获取该食物的过程不会被人类察觉,这时它们会选择获取那块被遮挡的食物。在上述两种条件下,如果看不到人类,它们同样会选择获取食物(所有条件下,黑猩猩都躲在一个障碍物的后面,需要通过一个透明或不透明管状容器获取食物)。最让人印象深刻的是,在获取食物的过程中,黑猩猩会采用安静的方式悄悄靠近食物,尽量不引起人类实验者的注意。上面这些不同的实验结果显示出,黑猩猩在需要使用完全不同知觉模型的问题情境中表现出泛化的处理能力。这也进一步提示,其间涉及的认知模型和推理具有很强的灵活性和生态效度。
同物理认知相比,上述研究中黑猩猩不仅能够基于对意图的一般理解进行很多相关推理,同时可以把这些推理镶嵌到一个原型结构之中,对推理对象的行为进行预测甚至操控(见图2-3)。
这些实验中与个体获取食物相关的背景信息是,竞争对手在且只有在具有食物获取目标同时意识到食物的位置时才会选择获取食物。黑尔等人(Hare et al.,2000)的实验中黑猩猩表现出的原型条件性推理直接反映了上述内容:如果居于支配地位的黑猩猩想吃香蕉同时看到香蕉在A处,那么它会直接去A处拿香蕉。类比类人猿的物理性认知,如果我们同样将原型否定当作包含互斥构念的一个连续体,上述食物竞争实验中黑猩猩亦表现出了原型否定的使用。黑尔等人实验中的黑猩猩懂得,如果竞争对手仅仅看到了障碍物,那么它会待在原地(如果看不到食物就不会想要获取它,见图2-3中的C条件)。梅利斯等人(Melis,2006a)隐藏实验中的黑猩猩则知道,如果实验者仅仅看到障碍物或摇晃杯子时听不到声响,那么她就会静静地坐着(如果看不到也听不到我靠近食物,她就不会想要获取食物),这也意味着只有在另一个个体看到障碍物或听到“安静”时,黑猩猩才会去拿食物[4]。

图2-3 食物竞争条件下的类人猿推理
总的看来,如同物理认知中的表现,类人猿在社会认知中亦很擅长操控。这种特殊技能在它们之间的肢体语言交流中同样能够体现出来(它们的发声通信与猴类相似,大都是固定化的,所以对思维问题研究的价值不大)。四种类人猿都可以通过特殊的(非猿灵长类大多不具有)姿势语言进行交流。经过彼此之间不断的社会互动,它们对一些意图动作逐渐仪式化。例如,把举起手臂进行击打当作邀请同类游戏的信号,并灵活地利用此类形式对其他个体的行为进行操控。更为重要的是,它们可以使用一系列吸引注意的姿势语言,以达到操控其他个体注意力的目的,如通过拍打地面吸引同类的注意。此外,它们甚至能够采用人类的“够”“指”等姿势性动作——类人猿在与同类沟通中本来并不存在的姿势语言——这清楚地显示出类人猿在操控其他个体行为及注意时具有的灵活性(Call & Tomasello,2007)。这样,类人猿的肢体沟通再一次显示出它们在操控原因时具有的独特技能。
关于类人猿对社交世界的理解,我们要介绍的最后一个实验同它们是否具有逆向推理有关。巴特尔曼等人(Buttelmann,2007)采用理性模仿范式(Gergely et al.,2002)对6只喂养的黑猩猩进行测试。实验中黑猩猩会看到实验者借助一个装置表演一些非常规动作,同时产生一些很有趣的结果。在一种条件下,情境中的物理限制会迫使实验者发出一些特殊动作,如由于手中拿着毛毯不得不用头来开灯、由于手中拿着一摞书不得不用脚激活音乐盒。实验者表演结束后,黑猩猩也被给予同样的装置,不同的是这时外界相关的物理限制被撤销了,结果发现,黑猩猩像往常一样发出动作,并不会像实验者那样采用一些非常规动作;然而当黑猩猩看到实验者在没有外界物理限制条件下采用一些非常规动作时,即便我们认为并不存在需要模仿的显性原因,它仍会进行模仿,如用头开灯。我们认为,对黑猩猩具有的这种区别性模仿最自然的解释就是它们使用了假言推理(proto-madus tollens),牵涉到原型否定的从结果到原因的回溯过程,这同卡尔(Call,2004)摇晃杯子研究中涉及的思维过程很相似,即(1)他没有使用他的双手;(2)如果让他自由选择,他会使用双手;(3)所以他的行为一定不是自由选择。
同解决复杂物理性问题相似,上述研究也提示,通过已有认知模型(对意图的一般理解)同化问题情境中的核心成分,之后运用该模型模拟并推理之前发生过什么或将来将会发生什么,类人猿同样能够解决很多复杂的社会性问题。同时在社会性推理的原逻辑范式框架下,类人猿在推理时可以运用原型条件性和原型否定(包含正向或逆向推理模式)。作为对本小节的总结,我们认为,与对物理世界的认知相似,在对社交世界的认知上,类人猿在上面提到的一些实验中的表现同样可以归为“思维”。
前面研究中提到的类人猿在相关实验中的表现,很明显不是简单的自动化行为反应或目标性反应,它包含着某种服务于更优决策的监控机制。行为层面,近来研究已经发现类人猿可以:
(1)为了未来更大的奖赏而延迟获得即时奖励;
(2)为了成功应对新的问题情境而抑制已有的相关问题解决策略;
(3)为了获得心仪的奖励而做一些自己不喜欢做的事;
(4)失败后的坚持;
(5)在易分散注意力情境下保持注意力集中。
此外,在一个系统性比较研究中,研究者发现黑猩猩在上述能力上的表现同3岁的人类婴儿水平相当(Herrmann et al.,submitted[5])。实际上,上述提及的黑猩猩具有的相关能力都有固定术语,即冲动抑制、注意控制、情绪调节、执行功能。然而在这里,我们把那些基于行为的自我调节称为行为自我监控(behavioral self-monitoring);把那些涉及的认知部分称为认知性自我监控(cognitive seif-monitoring)。
类人猿不仅具有行为自我监控能力,同时还可以进行认知性自我监控,这已被诸多同时采用多种不同范式的非人灵长类研究所证实。在最经典的范式中(通常以恒河猴为被试),个体为了获得心仪的奖赏必须学会区分(或记住一些东西),如果它们不能正确区分(或记忆),就不能获得奖赏并且必须在下一个试次前暂停一段时间。其中实验涉及的一个重要操作是,每一个试次中,个体都可以选择退出任务(意味着可以立即进入下一个试次),同时100%获得一个相对较小的奖励。这样,对于那些很可能区分或记忆失败的任务,个体逐渐学会了采用退出策略(Hampton,2001),它们似乎知道自己不知道什么或者记不住什么。
在涉及黑猩猩的另一个常用范式中,个体可以/不可以看到食物被隐藏进某个管状容器的过程,结果发现,当黑猩猩看到食物的隐藏过程时,它们会直接到对应的管状容器中寻找;看不到时,个体不知道食物被藏到哪里,选择时就会有一定的困难,这种情况下,黑猩猩似乎知道自己不知道相关信息,或者至少对相关信息不确定,就会试图做些什么。很有趣的是,这个过程中涉及的相关变量同样表现在人类身上:如果奖赏具有很大的诱惑性,或者距离获得相关信息已经比较久,个体就会有更强的寻找相关信息的趋向(Call,2010)。总的来说,当类人猿在评估情境以决定做什么的时候,如有没有充足的相关信息,它们会主动搜寻信息以服务于决策需要。
当然,对前面提到的一些实验的解释可能有很多种,但至少我们可以确定的是,类人猿可以进行某些类别的自我监控和自我评估。同时值得注意的是,它们不仅可以监控想象中的行为和后果/原因与结果,也可以监控自己的知识和记忆,并通过这种监控对某种行为成功的可能性进行估计。也就是说,至少在功用性背景下,类人猿和其他灵长类动物是可以对自身心理状态进行评估的,虽然这种评估不能等同于人类的自我反思(同后者相比,前者缺少社会/前景维度),但它至少为类人猿具有可以被称为思维的三个核心成分(抽象性认知表征、原逻辑推理范式、心理的自我监控和评估)提供了进一步的证据支持。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










