



三、物质循环
生态系统中的物质主要指生物维持生命活动正常进行所必需的各种营养元素。包括近30种化学元素,其中主要的是碳、氢、氧、氮和磷5种,它们构成全部原生质的97%以上。这些营养物质存在于大气、水域及土壤中。
生态系统中各种营养物质经过分解者分解成可被生产者利用的形式归还环境中重复利用,周而复始地循环,这个过程叫物质循环。物质通过食物链各营养级传递和转化,完成生态系统的物质流动。
(一)生态系统物质循环的层次及类型
1.生态系统物质循环的层次
(1)生物个体层次的物质循环。在这个层次上,生物个体吸收营养物质建造自身,经过代谢活动,生物从外界取得生存必需的物质,并使这些物质变成生物本身的物质,同时把体内产生的废物排出体外。这种新物质代替旧物质的过程叫做新陈代谢(简称代谢),通过代谢作用又把物质排出体外,经过分解者的作用归还于环境。
(2)生态系统层次(生态系统内)的物质循环。在初级生产者的代谢基础上,通过各级消费者和分解者把营养物质归还环境之中,又称为生物小循环或营养物质循环。这一循环是在一个具体范围内进行的(某一生态系统内),物质循环流速快、周期短。
(3)生物圈层次的物质循环(生物地球化学循环)。这一层次的物质循环是营养物质在各生态系统之间的输入与输出,以及它们在大气圈、水圈和土壤圈之间的交换。“生物地球化学循环”又称“生物地质化学循环”。因为生物体全部原生质约有97%以上由氧、碳、氢、氮、磷五种元素组成,它们在生物圈中的物质循环过程分属生物、地质、化学系统。这些营养物质存在于大气、水域及土壤中。如果说,生态系统能量的来源是太阳,那么,物质的来源便是生物栖身的地球,即地球上的大气圈、水圈、岩石圈及土壤圈。一个来自“天”,一个来自“地”,正是这“天”与“地”的结合,才有了生命所需要的能量和物质。
2.生态系统物质循环的类型
生物地球化学循环包括地质大循环和生物小循环两部分内容。根据物质在循环时所经历的路径的不同,从整个生物圈的观点出发,生物地球化学循环可分为气态循环型和沉积循环型两类。
(1)地质大循环和生物小循环:
①地质大循环。物质或元素经生物体的吸收作用,从环境进入生物有机体内,生物有机体再以死体、残体或排泄形式将物质或元素返回环境,进入大气、水、岩石、土壤和生物五大自然圈层的循环。地质大循环时间长、范围广,是闭合式循环(见图13-10)。
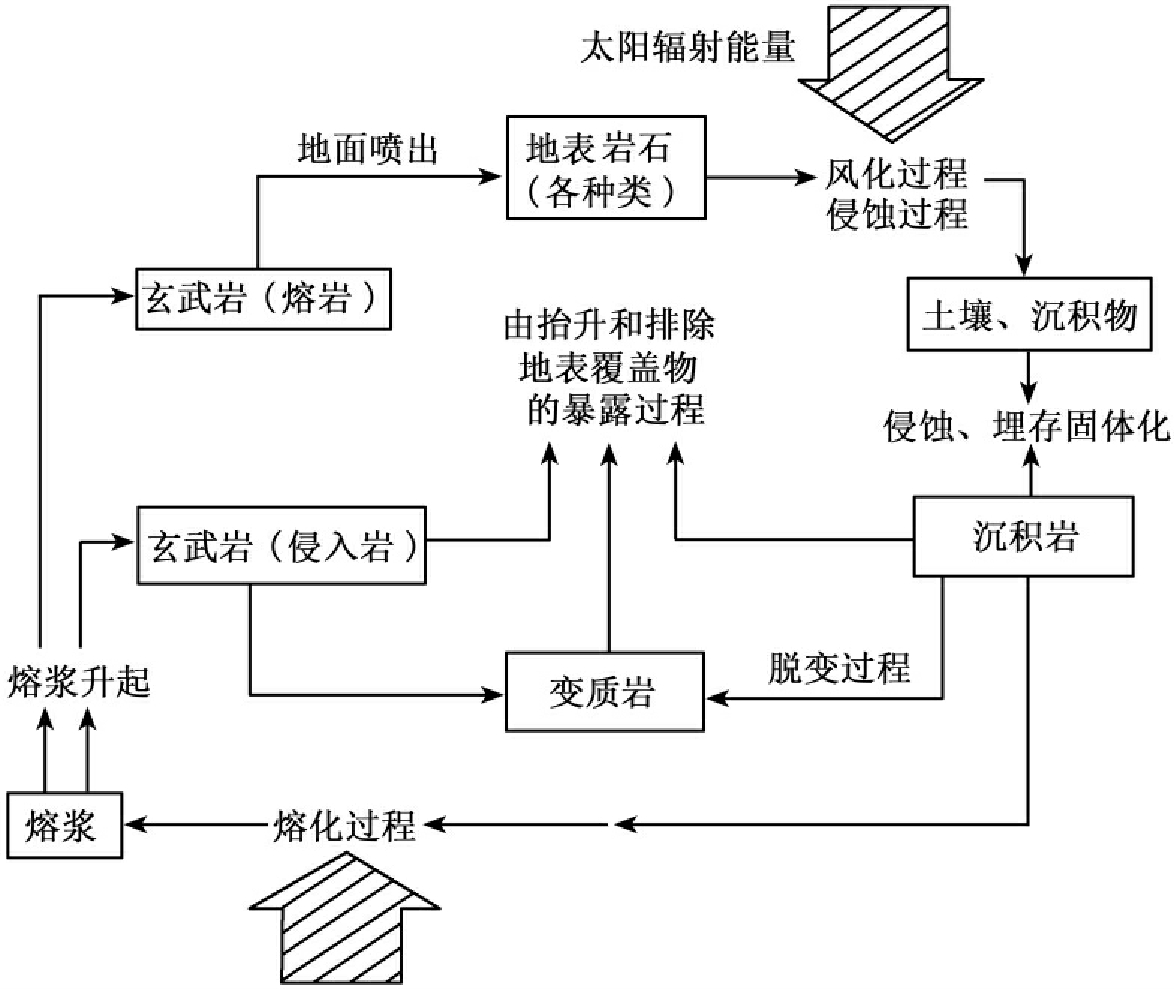
图13-10 地质循环的主要过程(据Copyright,1977)
②生物小循环。环境中的元素经生物体吸收,在生态系统中被多层次利用,然后经过分解者的作用,再为生产者吸收、利用。生物小循环的时间短、范围小,是开放式的循环(见图13-11)。
地质大循环是物质通过四大自然圈层的循环,具有全球性质,循环周期较长。如整个大气圈中的CO2,通过生物圈中生物的光合作用和呼吸作用,约300年循环一次;O2通过生物代谢约2000年循环一次;水圈中的水(包括占地球表面71%的海洋)通过生物圈
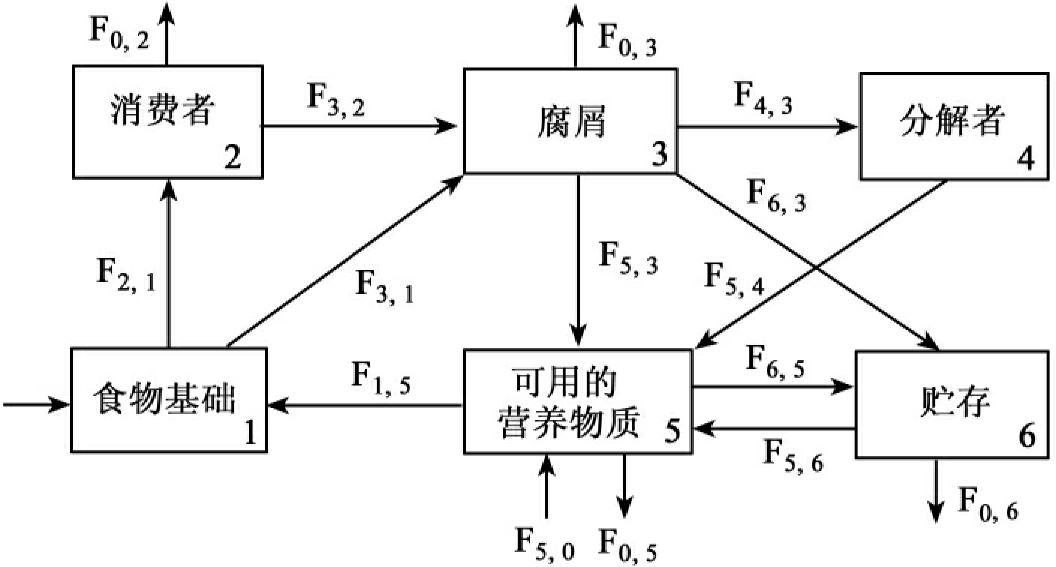
箭头表示营养物质流向(F),方块中数字表示分室号数,O代表环境; F2,1表示物质从1分室流向2分室
图13-11 营养物质生物地化循环
的吸收、排泄、蒸发、蒸腾,约200万年循环一次。至于由岩石、土壤风化出来的矿物元素,循环一次则需要更长时间,有的可长达几亿年。而生物小循环则要快得多,如水10天就可以循环一次。
(2)气相型循环和沉积型循环
①气相型循环(Gaseous Type Cycles)。其贮存库在大气圈或水圈(海洋)中,即元素或化合物可以转化为气体形式,通过大气进行扩散,弥漫在陆地或海洋上空,在很短时间内可以为植物重新利用,循环比较迅速,例如碳、氮、氧等。由于有巨大的大气贮存库,对于干扰可相当快地进行自我调节(但大气的自我调节也不是无限的)。因此,从全球意义上看,这类循环是比较完全的循环。
②沉积型循环(Sedimentary Type Cycles)。许多矿物元素其贮存库在地壳里,经过自然风化和人类的开采冶炼,从陆地岩石中释放出来,为植物所吸收,参与生命物质的形成,并沿食物链转移。然后动植物残体或排泄物经微生物的分解作用,将元素返回环境。除一部分保留在土壤中供植物吸收利用外,一部分以溶液或沉积物状态随流水进入江河,汇入海洋,经过沉降、淀积和成岩作用变成岩石,当岩石被抬升并遭受风化作用时,该循环才算完成。这类循环是缓慢的,并且容易受到干扰,成为“不完全”的循环,受到生物作用的负反馈调节,变化较小。
(二)物质循环特征
1.库(Pool)与流(Flow)
物质在运动过程中被暂时固定、贮存的场所称为库。生态系统中的各个组分都是物质循环的库,可分为植物库、动物库、大气库、土壤库和水体库。各库又可分为许多亚库,如植物库可分为作物、林木、牧草等亚库。在生物地球化学循环中,物质循环的库可归为两大类:一为贮存库,其容积较大,物质交换活动缓慢,一般为非生物成分的环境库;二为交换库,其容积较小,与外界物质交换活跃,一般为生物成分。例如,在一个水生生态系统中,水体中含有磷,水体是磷的贮存库;浮游生物体内含有磷,浮游生物是磷的交换库。
物质在库与库之间的转移运动状态称为流。生态系统中的能流、物流和信息流,使生态系统各组分密切联系起来,并使系统与外界环境联系起来。没有库,环境资源不能被吸收、固定、转化为各种产物;没有流,库与库之间不能联系、沟通,则物质循环短路,生命无以维持,生态系统必将瓦解。
2.生物量与现存量
在某一特定观察时刻,单位面积或体积内积存的有机物质总量称为生物量。它可以是特指的某种生物的生物量,也可以指全部植物、动物和微生物的生物量。多数人又将生物量称为现存量。生产量是现存量与减少量之和。减少量是指由于被取食、寄生或死亡、脱毛、产茧等损失的量,不包括呼吸损失量。生产量高的生态系统,生物现存量不一定大,例如,某生态系统的生产量为500kg,由于减少量为零,则现存量为500kg,另一生态系统的生产量为5000kg,但由于减少量为4500kg,其现存量也只有500kg。在生态研究中通常测定的是现存量及由其推算的净生产量。
3.周转率与周转期
周转率和周转期是衡量物质流动(或交换)效率高低的两个重要指标。周转率(R)是指系统达到稳定状态后,某一个组分(库)中的物质在单位时间内所流出的量(F0)或流入的量(F1)占库存总量(S)的份额。周转期是周转率的倒数,表示该组分的物质全部更换平均需要的时间。
周转率(R)=F1/S=F0/S
周转期(T)=1/周转率=1/R
物质在运动过程中,周转速率越高,则周转一次所需时间越短。
物质的周转率用于生物的生长称为更新率。某段时间末期,生物的现存量相当于库存量(S);在该段时间内,生物的生长量(P)相当于物质的输出量(F0)。不同生物的更新率相差悬殊,一年生植物当生育期结束时生物的最大现存量与年生长量大体相等,更新率接近1,更新期为1年。森林的现存量是经过几十年甚至几百年积累起来的,所以比年净生产量大得多,如某一森林的现存量为324t/hm2,年净生产量为28.6t/hm2,其更新率=28.6/324=0.088,更新期约11.3年。至于浮游生物,由于其代谢率高,现存生物量常常是很低的,但却有着较高的年生产量,如某一水体中浮游生物的现存量为0.07t/ hm2,年净生产量为4.1t/hm2,其更新率=4.1/0.07=59,更新期只有6.23天。
4.循环效率
当生态系统中某一组分的库存物质,一部分或全部流出该组分,但并未离开系统,并最终返回该组分时,系统内发生了物质循环。循环物质(FC)占总输入物质(F1)的比例,称为物质的循环效率(EC)。
EC=FC/F1
5.生态系统内能流与物流的关系
生态系统内,同时存在着能流与物流,它们相伴而行、相辅相成,且不可分割。能流是物流的动力,物流是能流的载体。物质的循环过程,是物质由简单无机态到复杂有机态再回到简单无机态的再生过程,同时也是系统的能量由生物固定、转化和消散的过程。物质也好,能量也好,不管它们的形态发生怎样的变化,都遵循着守恒的原则。但是,相对于生态系统而言,由于太阳能为主要能源,是无限的,而物质却是有限的,分布也是很不均匀的。
进一步比较流经生态系统的能流和物流,它们之间还有很多的区别,如能流单向流动并且在转化过程中逐渐衰变,有效能的数量逐级减少,最终趋向于全部转化为低效热能,离开生态系统。生态系统中某些贮存的能量,也能形成逆向的反馈能流,但能量只能被利用一次,所谓再利用是指未被利用过的部分。而物流不是单方向流动,而是往复循环,不是只利用一次,而是重复利用,物质在流动的过程中只是改变形态而不会消灭,可以在系统内永恒地循环,不会成为废物。
任何生态系统的存在和发展,都是能流与物流同时作用的结果,二者有一方受阻都会危及生态系统的延续和存在。
6.物质循环的调节
生态系统物质循环的自我调节作用表现在多方面,循环中每一个库和流因外来干扰引起的变化,都会引起有关生物的相应变化,产生负反馈调节使变化趋向消失而恢复稳态。大气中二氧化碳浓度上升会使光合作用增强;土壤中有效氮的缺乏,使共生、自生固氮微生物大量发展;水域富营养化水藻和水生植物恶性繁殖等都是这种负反馈作用的例子。
人对生态系统物质循环的干预,如果忽视了生态系统固有的自我调节能力,就有可能使某些有益的负反馈机制削弱或破坏,而导致系统发展失控直至产生严重的恶果。目前人类盲目地破坏植被、乱捕滥杀某些动物、大量地不适当地施用各种化学品以及向生态系统排放各类未经处理的污染物质所产生的后果都是这方面的例子。
(三)典型物质循环过程
1.碳循环
碳是一切生物体中最基本的成分,有机体干重的45%以上是碳。
据估计,全球碳贮存量约为26×1015t,但绝大部分以碳酸盐的形式禁锢在岩石中,其次是贮存在化石燃料中。生物可直接利用的碳是水圈和大气圈中以二氧化碳形式存在的碳,二氧化碳或存在于大气中或溶解于水中,所有生命的碳源均是二氧化碳。碳的主要循环形式是从大气的二氧化碳蓄库开始,经过生产者的光合作用,把碳固定,生成糖类,然后经过消费者和分解者,在呼吸和残体腐败分解后,再回到大气蓄库中。碳被固定后始终与能流密切结合在一起,生态系统的生产力的高低也是以单位面积中碳来衡量的。
植物通过光合作用,将大气中的二氧化碳固定在有机物中,包括合成多糖、脂肪和蛋白质,而贮存于植物体内。食草动物吃了以后经消化合成,通过一个一个营养级,再消化再合成。在这个过程中,部分碳又通过呼吸作用回到大气中;另一部分成为动物体的组分,动物排泄物和动植物残体中的碳,则由微生物分解为二氧化碳,再回到大气中。
除了大气,碳的另一个储存库是海洋,它的含碳量是大气的50倍,更重要的是海洋对于调节大气中的含碳量起着重要的作用。在水体中,同样由水生植物将大气中扩散到水上层的二氧化碳固定转化为糖类,通过食物链经消化合成,再消化再合成,各种水生动植物呼吸作用又释放二氧化碳到大气中。动植物残体埋入水底,其中的碳都暂时离开循环,但是经过地质年代,又可以石灰岩或珊瑚礁的形式再出露于地表;岩石圈中的碳也可以借助于岩石的风化和溶解、火山爆发等重返大气圈,有部分则转化为化石燃料,燃烧过程使大气中的二氧化碳含量增加(见图13-12)。
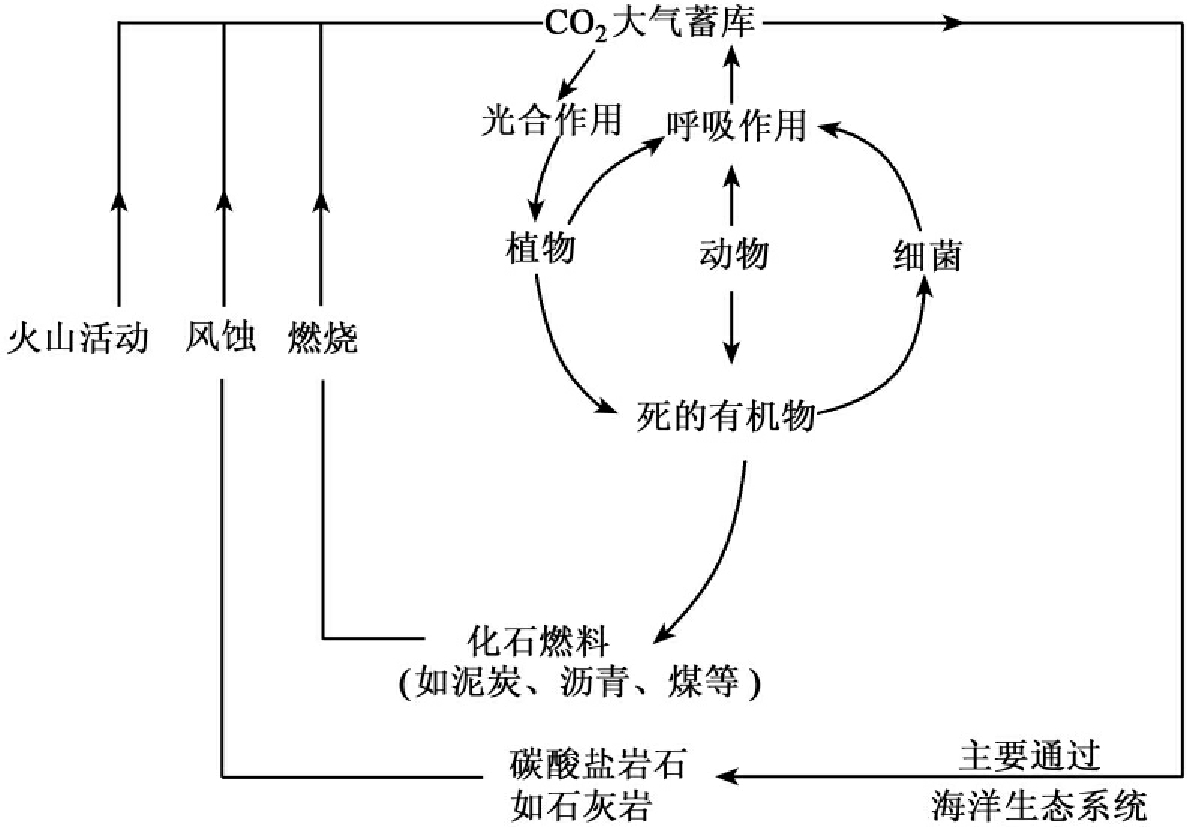
图13-12 生态系统中的碳循环
自然生态系统中,植物通过光合作用从大气中摄取碳的速率与通过呼吸和分解作用而把碳释放到大气中的速率大体相同。由于植物的光合作用和生物的呼吸作用受到很多地理因素和其他因素的影响,所以大气中的二氧化碳含量有着明显的日变化和季节变化。例如,夜晚由于生物的呼吸作用,可使地面附近的二氧化碳的含量上升,而白天由于植物在光合作用中大量吸收二氧化碳,可使大气中二氧化碳含量降到平均水平以下;夏季植物的光合作用强烈,因此,从大气中所摄取的二氧化碳超过了在呼吸和分解过程中所释放的二氧化碳,冬季正好相反,其浓度差可达0.002%。
二氧化碳在大气圈和水圈之间的界面上通过扩散作用而相互交换。二氧化碳的移动方向,主要决定于在界面两侧的相对浓度,它总是从高浓度的一侧向低浓度的一侧扩散。借助于降水过程,二氧化碳也可进入水体。1L雨水中大约含有0.3mL的二氧化碳。在土壤和水域生态系统中,溶解的二氧化碳可以和水结合形成碳酸,这个反应是可逆的,反应进行的方向取决于参加反应的各成分的浓度。碳酸可以形成氢离子和碳酸氢根离子,而后者又可以进一步离解为氢离子和碳酸根离子。由此可以预见,如果大气中的二氧化碳发生局部短缺,就会引起一系列的补偿反应,水圈中的二氧化碳就会更多地进入大气圈中;同样,如果水圈中的二氧化碳在光合作用中被植物利用耗尽,也可以通过其他途径或从大气中得到补偿。总之,碳在生态系统中的含量过高或过低都能通过碳循环的自我调节机制而得到调整,并恢复到原有水平。大气中每年大约有1×1011t的二氧化碳进入水体,同时水中每年也有相同数量的二氧化碳进入大气中,在陆地和大气之间,碳的交换也是平衡的,陆地的光合作用每年大约从大气中吸收1.5×1010t碳,植物死后被分解约可释放出1.7×1010t碳,森林是碳的主要吸收者,每年约可吸收3.9×109t碳。因此,森林也是生物碳的主要贮库,约储存482×109t碳,这相当于目前地球大气中含碳量的2/3。
在生态系统中,碳循环的速度是很快的,最快的在几分钟或几小时就能够返回大气,一般会在几周或几个月返回大气。一般来说,大气中二氧化碳的浓度基本上是恒定的。但是,近百年来,由于人类活动对碳循环的影响,一方面森林大量砍伐,同时在工业发展中大量化石燃料的燃烧,使得大气中二氧化碳的含量呈上升趋势。由于二氧化碳对来自太阳的短波辐射有高度的透过性,而对地球反射出来的长波辐射有高度的吸收性,这就有可能导致大气层低处的对流层变暖,而高处的平流层变冷,这一现象称为温室效应。由温室效应而导致地球气温逐渐上升,引起未来的全球性气候改变,促使南北极冰雪融化,使海平面上升,将会淹没许多沿海城市和广大陆地。虽然二氧化碳对地球气温影响问题还有很多不明之处,有待人们进一步研究,但大气中二氧化碳浓度不断增大,对地球上生物具有不可忽视的影响,这一点是毋庸置疑的。
2.氮循环
氮是蛋白质的基本成分,因此,是一切生命结构的原料。
虽然大气化学成分中氮的含量非常丰富,有78%为氮,然而氮是一种惰性气体,植物不能够直接利用。因此,大气中的氮对生态系统来讲,不是决定性库。必须通过固氮作用将游离氮与氧结合成为硝酸盐或亚硝酸盐,或与氢结合成氨,才能为大部分生物所利用,参与蛋白质的合成。因此,氮被固定后,才能进入生态系统,参与循环。
固氮的途径有三种。一是通过闪电、宇宙射线、陨石、火山爆发活动的高能固氮,其结果形成氨或硝酸盐,随着降雨到达地球表面。据估计,通过高能固定的氮大约为8.9kg/(hm2·a)。二是工业固氮,这种固氮形式的能力已越来越大。20世纪80年代初全世界工业固氮能力已为3×107t,到20世纪末,已接近1×108t。第三条途径,也是最重要的途径就是生物固氮,为100~200kg/(hm2·a),约占地球固氮的90%。能够进行固氮的生物主要是固氮菌,与豆科植物共生的根瘤菌和蓝藻等自养和异养微生物。在潮湿的热带雨林中生长在树叶和附着在植物体上的藻类和细菌也能固定相当数量的氮,其中一部分固定的氮为植物本身所利用。
氮在环境中的循环可用图13-13来表示。植物从土壤中吸收无机态的氮,主要是硝酸盐,用做合成蛋白质的原料。这样,环境中的氮进入了生态系统。植物中的氮一部分为草食动物所取食,合成动物蛋白质。在动物代谢过程中,一部分蛋白质分解为含氮的排泄物(尿素、尿酸),再经过细菌的作用,分解释放出氮。动植物死亡后经微生物等分解者的分解作用,使有机态氮转化为无机态氮,形成硝酸盐。硝酸盐可再为植物所利用,继续参与循环,也可被反硝化细菌作用,形成氮气,返回大气库中。
因此,含氮有机物的转化和分解过程主要包括有氨化作用、硝化作用和反硝化作用。
氨化作用——由氨化细菌和真菌的作用将有机氮(氨基酸和核酸)分解成为氨与氨化合物,氨溶于水即成为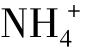 ,可为植物所直接利用。
,可为植物所直接利用。
硝化作用——在通气情况良好的土壤中,氨化合物被亚硝酸盐细菌和硝酸盐细菌氧化为亚硝酸盐和硝酸盐,供植物吸收利用。土壤中还有一部分硝酸盐变为腐殖质的成分,或被雨水冲洗掉,然后经径流到达湖泊和河流,最后到达海洋,为水生生物所利用。海洋中还有相当数量的氨沉积于深海而暂时离开循环。
反硝化作用——也称脱氮作用,反硝化细菌将亚硝酸盐转变为大气氮,回到大气库中。
因此,在自然生态系统中,一方面通过各种固氮作用使氮素进入物质循环,而通过反硝化作用、淋溶沉积等作用使氮素不断重返大气,从而使氮的循环处于一种平衡状态。
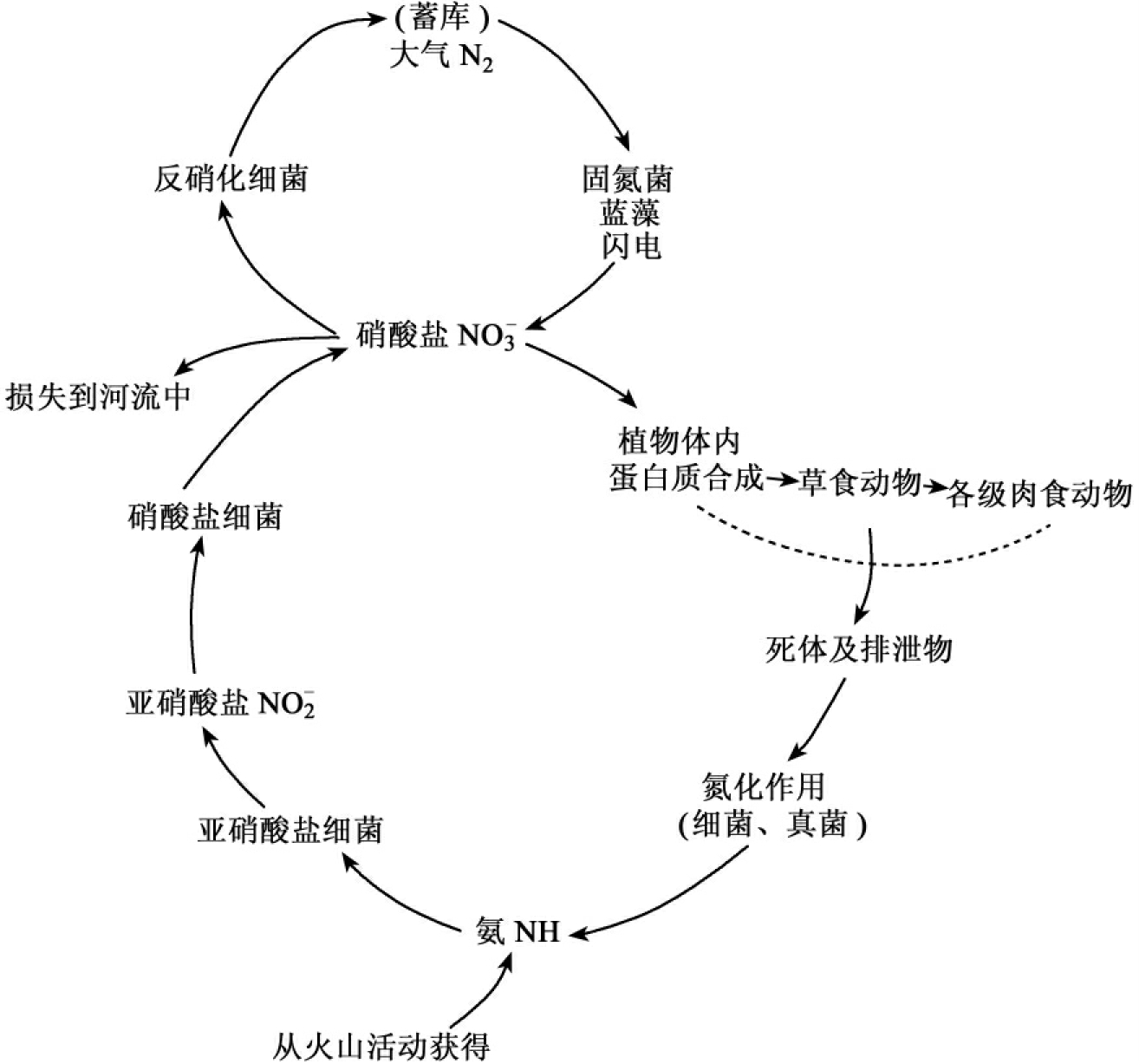
图13-13 生态系统中的氮循环
3.磷循环
磷是生物不可缺少的重要元素,生物的代谢过程都需要磷的参与,磷是核酸、细胞膜和骨骼的主要成分,高能磷酸键在腺苷二磷酸(ADP)和腺苷三磷酸(ATP)之间可逆地转移,它是细胞内一切生化作用的能量。
磷不存在任何气体形式的化合物,所以磷是典型的沉积型循环物质。沉积型循环物质主要有两种存在相:岩石相和溶解盐相。循环的起点源于岩石的风化,终于水中的沉积。由于风化侵蚀作用和人类的开采,磷被释放出来,由于降水成为可溶性磷酸盐,经由植物、草食动物和肉食动物而在生物之间流动,待生物死亡后被分解,又使其回到环境中。溶解性磷酸盐,也可随着水流进入江河湖海,并沉积在海底。其中一部分长期留在海里,另一些可形成新的地壳,在风化后再次进入循环(见图13-14)。
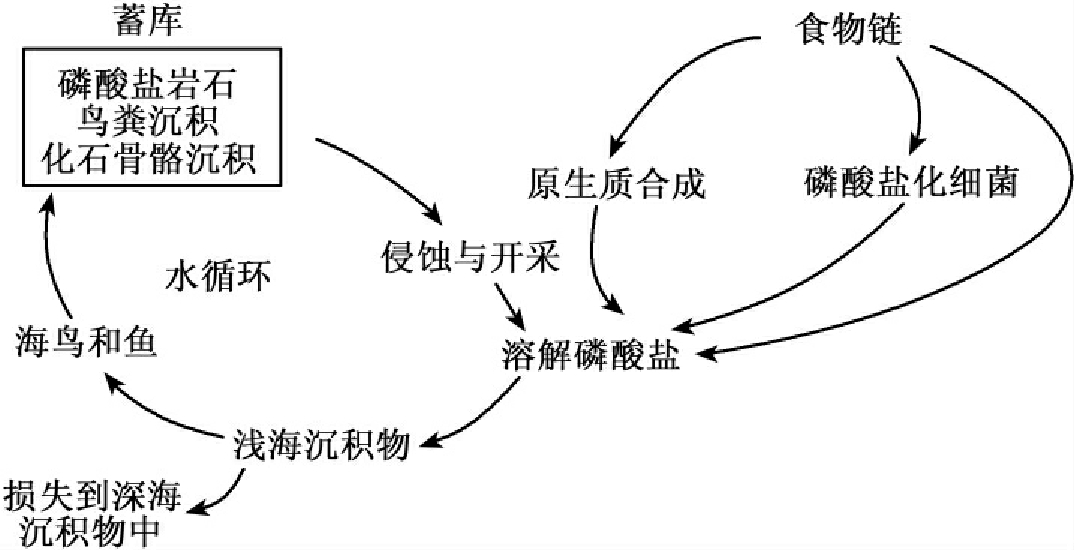
图13-14 生态系统中的磷循环
在陆地生态系统中,含磷有机物被细菌分解为磷酸盐,其中一部分又被植物再吸收,另一些则转化为不能被植物利用的化合物。同时,陆地的一部分磷由径流进入湖泊和海洋。在淡水和海洋生态系统中,磷酸盐能够迅速地被浮游植物所吸收,而后又转到浮游动物和其他动物体内,浮游动物每天排出的磷与其生物量所含有的磷相等,所以使磷循环得以继续进行。浮游动物所排出的磷又有一部分是无机磷酸盐,可以为植物所利用,水体中其他的有机磷酸盐可被细菌利用,细菌又被其他的一些小动物所食用。一部分磷沉积在海洋中,沉积的磷随着海水的上涌被带到光合作用带,并被植物所吸收。因动植物残体的下沉,常使得水表层的磷被耗尽而深水中的磷积累过多。磷是可溶性的,但由于磷没有挥发性,所以,除了鸟粪和对海鱼的捕捞,磷没有再次回到陆地的有效途径。在深海处的磷沉积,只有在发生海陆变迁,由海底变为陆地后,才有可能因风化而再次释放出磷,否则就将永远脱离循环。正是由于这个原因,使陆地的磷损失越来越大。因此,磷的循环为不完全循环,现存量越来越少,特别是随着工业的发展而大量开采磷矿加速了这种损失。据估计,全世界磷蕴藏量只能维持100a左右,在生物圈中,磷参与循环的数量,目前正在减少,磷将成为人类和陆地生物生命活动的限制因子。
4.硫循环
硫是原生质体的重要组分,它的主要蓄库是岩石圈,但它在大气圈中能自由移动,因此,硫循环有一个长期的沉积阶段和一个较短的气体阶段。在沉积相,硫被束缚在有机或无机沉积物中。
岩石库中的硫酸盐主要通过生物的分解和自然风化作用进入生态系统。
化能合成细菌能够在利用硫化物中含有的潜能的同时,通过氧化作用将沉积物中的硫化物转变为硫酸盐;这些硫酸盐一部分可以为植物直接利用,另一部分仍能生成硫酸盐和化石燃料中的无机硫,再次进入岩石蓄库中。
从岩石库中释放硫酸盐的另一个重要途径是侵蚀和风化,从岩石中释放出的无机硫由细菌作用还原为硫化物,土壤中的这些硫化物又被氧化成植物可利用的硫酸盐。
自然界中的火山爆发也可将岩石蓄库中的硫以硫化氢的形式释放到大气中,化石燃料的燃烧也将蓄库的硫以二氧化硫的形式释放到大气中,可为植物吸收。
硫循环与磷循环有类似之处,但硫循环要经过气体型阶段。
硫的主要蓄库是硫酸盐岩,但大气中也有少量的存在。虽然生物对硫的需要并不像对碳、氮和磷那么多,而且硫不会成为有机体生长的限制因子。但在硫循环中涉及许多微生物的活动,生物体需要硫合成蛋白质和维生素。植物所需要的大部分硫主要来自于土壤中的硫酸盐,同时可以从大气中的二氧化硫获得。植物中的硫通过食物链被动物所利用,或动植物死亡后,微生物对蛋白质的分解将硫释放到土壤中,然后再被微生物利用,以硫化氢或硫酸盐形式而释放硫。无色硫细菌既能将硫化氢还原为单质硫,又能将其氧化为硫酸;绿色硫细菌在有阳光时,能利用硫化氢作为氧接收者;生活于沼泽和河口的紫细菌能使硫化氢氧化,形成硫酸盐,进入再循环,或者被产生者生物所吸收,或为硫酸还原细菌所利用(见图13-15)。
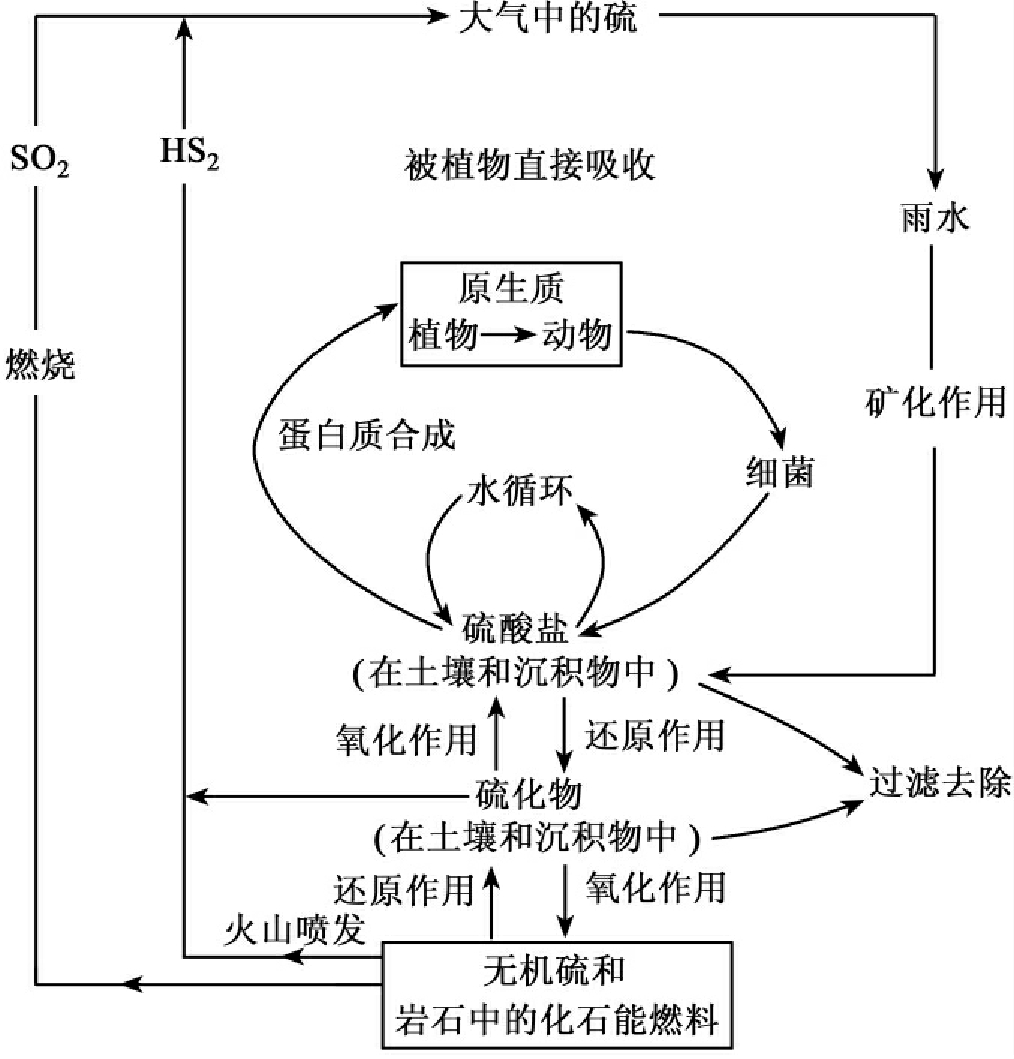
图13-15 生态系统中的硫循环
人类对硫循环的影响很大,通过燃烧化石燃料,人类每年向大气中输入的二氧化硫已达1.47×108t,其中70%来源于燃烧煤。二氧化硫在大气中遇水蒸气反应形成硫酸。大气中的硫酸对于环境有许多方面的影响,对人类及动物的呼吸道产生刺激作用,如果是细雾状的微小颗粒,还能进入肺,刺激敏感组织。二氧化硫浓度过高,就会成为灾害性的空气污染,例如伦敦1952年、纽约和东京1960年的二氧化硫灾害,造成支气管性哮喘大增,死亡率上升。空气中的污染物的种类很多,现在往往将硫的浓度作为空气污染严重程度的指标,空气中硫含量与人的健康关系最为密切。
(四)有毒有害物质循环与人体健康
1.有毒有害物质循环
全球化学污染是指化学物质在全球水平上对生物——非生物复合系统所具有的消极作用,致使它在全球规模上影响着生物的生存和人体的健康,它是化学物质生物地球化学循环引起的负面效应。也就是说,污染物质的生物地球化学循环是其遍布全球并进入食物链的基本方式。例如,PCB(多氯联苯)、二恶英和农药尽管在局部地区已禁止使用,但它们通过大气分室的生物地球化学循环,使世界范围内的大湖泊及浮游生物、鱼类中都含有这些化学物质。
有毒有害物质的循环是指那些对有机体有毒有害的物质进入生态系统,通过食物链富集或被分解的过程。由于工农业迅速发展,人类向环境中投放的化学物质与日俱增,从而使生物圈中的有毒有害物质的数量与种类相应增加,这些物质一经排放到环境中便立即参与生态系统的循环,它们像其他物质循环一样,在食物链营养级上进行循环流动。所不同的是大多数有毒物质,尤其是人工合成的大分子有机化合物和不可分解的重金属元素,在生物体内具有浓缩现象,在代谢过程中不能被排除,而被生物体同化,长期停留在生物体内,造成有机体中毒、死亡。Hg、Cd、As、Cr、Cu等重金属污染已成为人类所面临的严重环境问题之一。Hg污染引起的“水俣病”、Cd污染引起的“痛痛病”(又称“骨痛病”)等公害事件曾为世界所震惊。随后人们对这些有毒有害重金属元素在生物圈的迁移、转化及循环十分关注,并积累了较丰富的基础资料。化学农药的大量生产和使用也加剧了全球化学污染的严重性。当化学农药进入生物-非生物复合系统后,一般在土壤、水、大气和生物产生一定的毒害作用,并有可能对某些生物地球化学过程产生“毁灭性”的影响。农药生物地球化学循环的最终效应,可能是导致生物圈处于化学农药的完全暴露之下,带来一系列不良影响。
因此,有毒物质的生态系统循环与人类的关系最为密切,但又最为复杂。有毒物质循环的途径、在环境中的滞留时间、在有机体内浓缩的数量和速度,以及作用机制和对有机体影响的程度等都是十分重要的问题。
一般情况下,毒性物质进入环境,常常被空气和水稀释到无害的程度,以至无法用仪器检测。即使是这样,对食物链上有机体的毒害依然存在。因为小剂量毒物在生物体内经过长期的积累和浓集,也可以达到中毒致死的水平。同时,有毒物质在循环中经过空气流动及水的搬运以及在食物链上的流动,常常使有毒物质的毒性增加,进而造成中毒的过程复杂化。在自然界也存在着对毒性物质分解,减轻毒性的作用,例如放射性物质的半衰期,以及某些生物对有毒物质的分解和同化作用;相反,也有某些有毒物质经过生态系统的循环后使毒性增加,例如汞的生物甲基化等。
与大量元素相比较,尽管有毒有害物质的数量少,但随着人类对环境的影响越来越大,向环境中排放的物质的数量和种类仍在增加,它对生态系统各营养级的生物的影响也与日俱增,甚至已引起生态灾难,所以对有毒物质在生态系统中循环规律的研究已成为保护人类自身所必须。
2.地表元素迁移与人体健康
自然界中由于环境条件的不同,地表元素发生迁移,常造成一些元素在地表分布的不均。在一些生态系统分散流失,在另一些生态系统中又积累。这种生物地化循环常常导致某些生态系统中生命元素含量的异常,或不足,或过剩,会造成植物、动物乃至人类的疾病。俗话说“一方水土养一方人”就是这个道理。这类疾病常呈现区域性,故称“地方病”。据统计,人体除必需的大量元素外,还需要铁、锰、硼等14种微量元素,在正常情况下,这些元素在人体内处于相对平衡状态。一旦平衡的稳定状态遭到破坏,病变就会发生。微量元素在人体内含量虽少,但对保持人体健康和生物的生长发育却有重要的意义。
生物地球化学地方病大多数与微量元素有关,它主要是由化学环境异常引起的人体健康效应。人体从环境中摄入的元素数量超出或低于人体正常需要量,就会产生代谢失调,出现病态反应,影响健康。
引起地表化学元素区域分异的因子是多样而复杂的,一是化学元素本身的性质和内部结构;二是各区域自然地理条件和自然地理特征;三是人类活动类型和生活习惯与方式。
地方性甲状腺肿是由环境严重缺碘而引起的一种世界性地方病。主要集中分布于世界几个著名大山脉——安第斯山、喜马拉雅山、阿尔卑斯山和比利牛斯山,以及新西兰岛、新几内亚岛和非洲的马达加斯加岛等,其中以亚、非、拉地区流行最为严重。碘迁移能力很强,容易淋溶流失,因此世界上缺碘地区分布很广,主要是受地质地貌-气候条件的影响,山区半山区及沙丘、河流两岸冈地都能使碘容易淋失。碘异常的程度与地方性甲状腺肿患病率之间表现为一种概率统计相关关系,即碘的缺乏或过剩程度越严重,地方性甲状腺肿患病率就越高。
氟过多而造成的地方性氟中毒是由于长期饮用、食用当地高氟水或食物引起的一种慢性全身性骨骼系统疾病。轻者牙齿出现氟斑牙,重者出现氟骨症。人体主要从饮水和饮食中摄取氟,大部分来自饮用水。氟既是人体必需微量元素,又是中毒性元素。氟的化学性质活泼,是一种易迁移元素,在自然环境中的各种环境条件都会影响其迁移和累积,因此,氟在环境中的分布很不均匀,形成以氟不足为特征的龋齿高发区和以氟过剩为特征的地方性氟中毒区。
地方性砷中毒是指由原生环境引起的砷中毒。地方性砷中毒病是通过饮用水而致病,例如,日本、新西兰等国温泉、火山温泉地区,美国俄勒冈州、中国台湾的台南县、内蒙古的包头附近地区、新疆奎屯等地,都是比较典型的地方性砷中毒病区。患者表现为体弱、头晕、头痛、疲乏、失眠等非特异性中枢神经系统中毒症状。少数患者出现再生性障碍贫血、营养不良等,严重者表现为肢体无力、行动困难、运动失调。
大骨节病是一种原因不明的地方性骨关节病,病区分布具有明显的区域性。主要侵犯儿童和青年,其中7~15岁学龄儿童为易感人群。病人主要表现为关节疼痛,增粗变形,运行障碍,肌肉萎缩。重者发育障碍,短肢畸形,丧失劳动力。该病主要分布在我国从东北向西南走向的一条狭长地带内,即黑龙江、吉林、辽宁、内蒙古、河北、河南、山东、山西、陕西、甘肃、青海、四川、西藏和北京等地,此外在日本、朝鲜北部、俄罗斯远东和西伯利亚地区均有发生。有关大骨节病病因主要有生物地球化学说和食物性真菌毒素中毒学说两种,其环境特征与自然环境因素有密切关系。我国大骨节病病带可划分为四种生态环境类型:山地针阔混交林棕褐土生态环境类型;黄土高原落叶阔叶林黑垆土生态环境类型;平原湖相沉积生态环境类型和沙漠沼泽草炭沉积生态环境类型。在这四种生态环境类型中普遍存在低硒、高腐殖酸的环境特征。事实证明,在大骨节病区改造低硒环境,增加硒的摄入量能有效地控制大骨节病的发生。
克山病是一种以心肌坏死为主要症状的地方病,因最早发现于我国东北地区克山县而得名。患者发病急,死亡率高。克山病主要分布于我国黑龙江、吉林、辽宁、内蒙古、河北、河南、山东、山西、陕西、甘肃、宁夏、青海、四川、云南、湖北及西藏等地的某些地区。从东北至西南呈一宽带状分布。克山病病因至今未明,目前主要有生物病因学说和生物地球化学学说。多数学者认为克山病与自然环境水土因素密切相关,缺硒或处于低硒水平是该病带的典型环境特征。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。












