



第二节 糖在体内的代谢概况
糖的分解代谢途径主要有无氧氧化、有氧氧化及磷酸戊糖途径。
一、糖的无氧氧化
在机体缺氧(如剧烈运动)时,葡萄糖或糖原分解生成乳酸,并产生能量,这个过程称为糖的无氧氧化。在这个代谢过程中尤以红细胞和运动时的骨骼肌较为活跃,因与酵母的生醇发酵非常相似,故又称为糖酵解。参与糖酵解反应的一系列酶存在于细胞质中,因此糖酵解的全部反应过程均在细胞质中进行。
(一)糖酵解的反应过程
糖酵解的反应过程分为三个阶段:第一阶段是葡萄糖分解生成磷酸丙糖,即己糖碳链裂解成2分子磷酸甘油醛并耗能的阶段;第二阶段是2分子磷酸甘油醛转变成2分子丙酮酸并产能的阶段;第三阶段是2分子丙酮酸还原生成乳酸的阶段。
1.葡萄糖分解生成2分子磷酸甘油醛
(1)葡萄糖磷酸化生成6-磷酸葡萄糖:葡萄糖在己糖激酶催化下,由ATP提供磷酸基和能量,生成6-磷酸葡萄糖。
![]()
此反应是不可逆反应,反应过程中消耗ATP。己糖激酶是关键酶。所谓关键酶是指在代谢途径中,催化不可逆反应步骤、起着控制代谢途径运转速度快慢作用的酶,其活性受到变构剂和激素的调节。
糖原进行糖酵解时,非还原端的葡萄糖单位先进行磷酸分解生成1-磷酸葡萄糖,再经磷酸葡萄糖变位酶催化生成6-磷酸葡萄糖,此过程不消耗ATP。
6-磷酸葡萄糖是一个重要的中间代谢产物,是多条糖代谢途径(糖酵解、有氧氧化、磷酸戊糖途径,以及糖原合成、糖原分解、糖异生)的连接点。
(2)6-磷酸葡萄糖异构为6-磷酸果糖:此反应在磷酸己糖异构酶催化下进行,是一个醛-酮异构化反应。
![]()
(3)6-磷酸果糖磷酸化生成1,6-二磷酸果糖:催化此反应的酶是6-磷酸果糖激酶,这是糖酵解途径的第二次磷酸化反应,消耗ATP且需Mg2+参与,此反应不可逆。6-磷酸果糖激酶是关键酶。
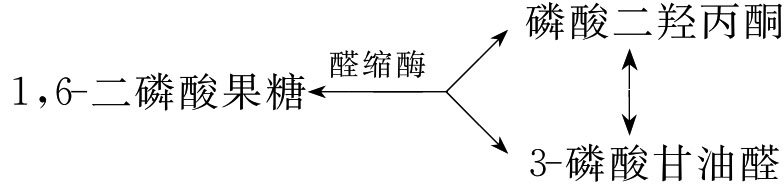
(4)1,6-二磷酸果糖裂解生成2分子的磷酸丙糖:此反应由醛缩酶催化,反应可逆。3-磷酸甘油醛和磷酸二羟丙酮互为异构体,在磷酸丙糖异构酶催化下可互相转变。当3-磷酸甘油醛在继续进行反应时,磷酸二羟丙酮可不断转变为3-磷酸甘油醛,这样1分子1,6-二磷酸果糖生成2分子3-磷酸甘油醛。
这一阶段的主要特点之一是葡萄糖的磷酸化。磷酸化后的化合物极性增高,不能自由进出细胞膜,因而葡萄糖磷酸化后不易逸出细胞外,反应限制在细胞质中进行;另外,结合了磷酸基团的化合物不仅能降低酶促反应的活化能,同时能提高酶促反应的特异性。
特点之二是伴随有能量的消耗。若从葡萄糖开始,消耗2分子ATP;若从糖原开始,则消耗1分子ATP。在这一阶段中有两个不可逆反应,分别由两个关键酶己糖激酶和6-磷酸果糖激酶催化,它们是糖酵解途径运转速度的调节点。
2.磷酸甘油醛氧化生成丙酮酸
(1)3-磷酸甘油醛氧化生成1,3-二磷酸甘油酸:在3-磷酸甘油醛脱氢酶的催化下,3-磷酸甘油醛脱氢并磷酸化,同时分子内部能量重排,生成含有高能磷酸键的1,3-二磷酸甘油酸,辅酶NAD+接受反应脱下的氢经还原生成NADH+H+。这是糖酵解中唯一的氧化反应。
![]()
(2)1,3-二磷酸甘油酸转变为3-磷酸甘油酸:1,3-二磷酸甘油酸在磷酸甘油酸激酶催化下将高能磷酸键转移给ADP生成ATP,自身转变为3-磷酸甘油酸。这是无氧酵解过程中第一次生成ATP。ATP的产生方式是底物水平磷酸化。此种由底物分子中的高能磷酸键直接转移给ADP而生成ATP的方式,称为底物水平磷酸化。
由于1分子葡萄糖能产生2分子1,3-二磷酸甘油酸,所以在这一过程中,1分子葡萄糖可产生2分子ATP。
![]()
(3)3-磷酸甘油酸转变为2-磷酸甘油酸:此反应由磷酸甘油酸变位酶催化,磷酸基团由3-位转至2-位。
![]()
(4)2-磷酸甘油酸脱水生成磷酸烯醇式丙酮酸:2-磷酸甘油酸经烯醇化酶催化进行脱水的同时,分子内部的能量重排,生成含有高能磷酸键的磷酸烯醇式丙酮酸。
![]()
(5)丙酮酸的生成:在丙酮酸激酶催化下,将磷酸烯醇式丙酮酸上的高能磷酸键转移给ADP生成ATP,自身则生成丙酮酸。这是糖酵解途径中的第二次底物水平磷酸化。此反应不可逆,其中,丙酮酸激酶是关键酶。由于1分子葡萄糖产生2分子丙酮酸,所以在这一过程中,1分子葡萄糖可产生2分子ATP。
![]()
第二阶段的特点是能量的产生。糖酵解过程的能量产生主要在3-磷酸甘油醛脱氢成为1,3-二磷酸甘油酸及磷酸烯醇式丙酮酸转变为丙酮酸的过程中,整个过程共产生4分子ATP,产生方式都是底物水平磷酸化。这一阶段中丙酮酸激酶是糖酵解过程的另一个关键酶和反应速度调节点。
3.丙酮酸还原生成乳酸
当机体缺氧时,在乳酸脱氢酶(LDH)催化下,由3-磷酸甘油醛脱氢反应生成的NADH+H+作为供氢体,将丙酮酸还原生成乳酸。NADH+H+重新转变成NAD+,糖酵解才能继续进行。
![]()
在有氧条件下,3-磷酸甘油醛脱氢产生的NADH+H+从细胞质进入线粒体经电子传递链传递生成水,同时释放出能量(详见第六章)。
在整个糖酵解的十步酶促反应中,有三步反应是不可逆的,催化这三步反应的酶——己糖激酶、6-磷酸果糖激酶、丙酮酸激酶是整个糖酵解过程的关键酶,调节这三个酶的活性可以影响糖酵解的运转速度。糖酵解的全过程如图7-2所示。
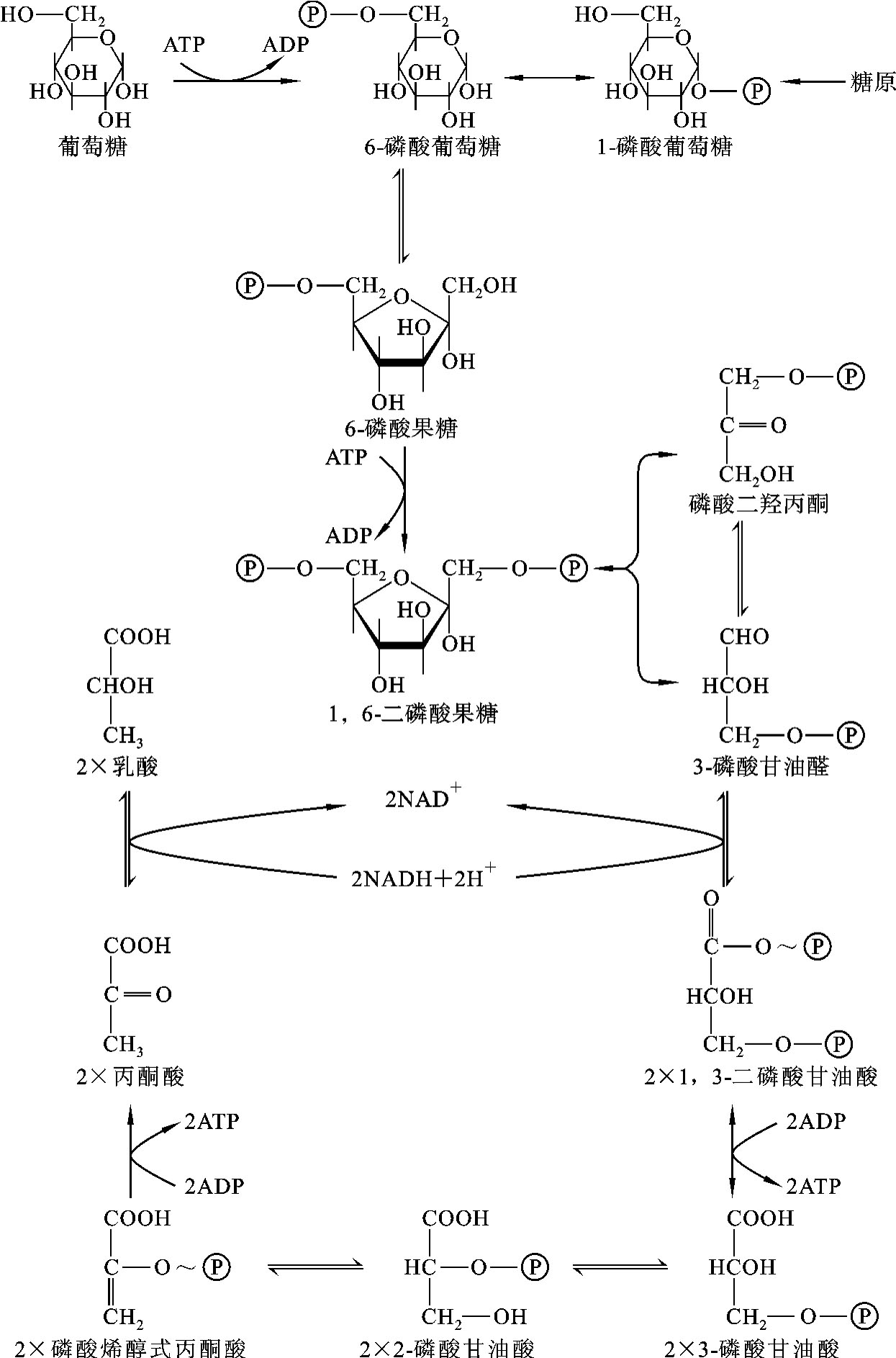
图7-2 糖酵解的全过程
(二)糖酵解的生理意义
1分子葡萄糖经糖酵解净生成2分子ATP;若从糖原开始,1分子葡萄糖单位净生成3分子ATP。糖酵解虽然产生的能量不多,但其生理意义特殊。
(1)迅速提供能量:这对肌肉组织尤为重要,肌肉组织中的ATP含量甚微,仅为5~7μmol/g(新鲜组织),肌肉收缩几秒钟就可全部耗尽。此时即使不缺氧,葡萄糖进行有氧氧化的过程也比糖酵解长得多,不能及时满足生理需要;而通过糖酵解则可迅速获得ATP。
(2)缺氧时的主要供能方式:如在剧烈运动时,肌肉局部血流不足会导致相对缺氧,必须通过糖酵解供能。某些病理情况,如严重贫血、大量失血、呼吸障碍、循环衰竭等,因供氧不足长时间依靠糖酵解供能,可导致乳酸堆积,而引起乳酸中毒。
(3)供氧充足时少数组织的能量来源:如视网膜、肾髓质、皮肤、睾丸等即便供氧充足,仍然依赖糖酵解供能。成熟红细胞没有线粒体,完全依靠糖酵解供能。此外,神经、白细胞、骨髓等代谢极为活跃,即使不缺氧,也常由糖酵解提供部分能量。
(4)糖酵解是糖有氧氧化的前段过程,其一些中间代谢物是脂类、氨基酸等合成的前体。
二、糖的有氧氧化
糖的有氧氧化是指葡萄糖或糖原在有氧条件下,彻底氧化分解生成CO2和H2O及能量的过程。机体绝大多数组织细胞能进行糖的有氧氧化。这是糖在体内氧化分解的主要方式,是机体获得能量的主要途径。
(一)有氧氧化的反应过程
糖的有氧氧化分为三个阶段:第一阶段是葡萄糖或糖原在细胞质中通过糖酵解途径分解生成丙酮酸;第二阶段是丙酮酸进入线粒体氧化脱羧生成乙酰CoA;第三阶段是乙酰CoA经三羧酸循环彻底氧化生成CO2、H2O和ATP。葡萄糖有氧氧化概况如图7-3所示。
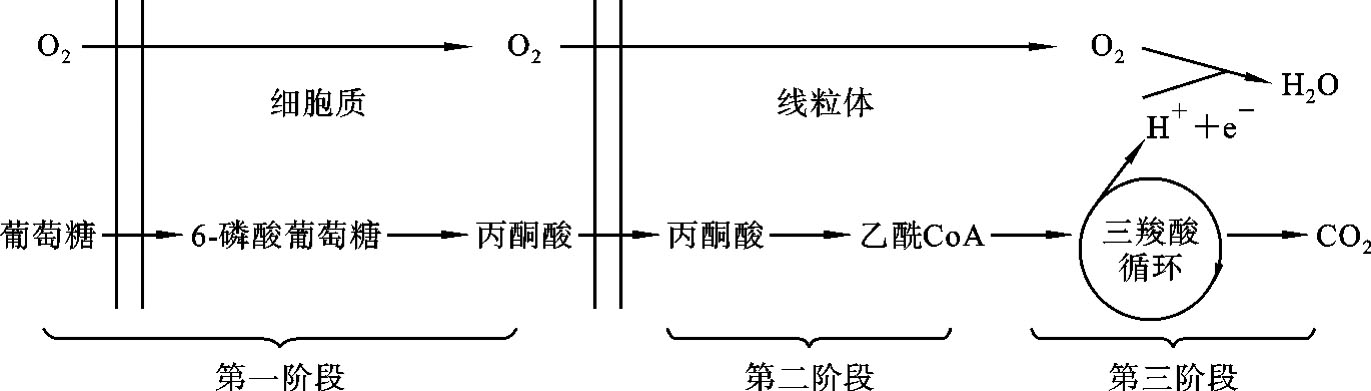
图7-3 葡萄糖有氧氧化概况
1.丙酮酸的生成
该反应与糖酵解途径相同,但反应中生成的NADH+H+不参与丙酮酸还原为乳酸的反应,而是经呼吸链氧化生成水并释放出能量。
2.乙酰CoA的生成
在细胞液中生成的丙酮酸在缺氧的条件下还原生成乳酸。在有氧的条件下丙酮酸则进入线粒体,然后在丙酮酸脱氢酶复合体的催化下,进行脱氢(氧化)和脱羧(脱去CO2),并与辅酶A(HSCoA)结合生成乙酰CoA。整个反应是不可逆的。
![]()
丙酮酸脱氢酶复合体是由丙酮酸脱氢酶、二氢硫辛酸转乙酰基酶、二氢硫辛酸脱氢酶三种酶组成的;该酶复合体需要多种含B族维生素的辅助因子,如TPP(含维生素B1)、HSCoA(含泛酸)、FAD(含维生素B2)、NAD+(含维生素PP)等。
多酶复合体中三种酶紧密相连,反应依次连锁迅速完成,催化效率高,使丙酮酸脱羧和脱氢生成乙酰CoA及NADH+H+。
3.三羧酸循环
由于三羧酸循环是Krebs于1937年发现的,故也称为Krebs循环。因循环以乙酰CoA与草酰乙酸缩合生成含有三个羧基的柠檬酸开始,经过一系列代谢反应,又生成草酰乙酸,故称为三羧酸循环(tricarboxylic acid cycle,TAC)或柠檬酸循环。
(1)柠檬酸的生成:乙酰CoA与草酰乙酸在柠檬酸合成酶催化下缩合生成柠檬酸。此反应不可逆,柠檬酸合成酶是关键酶。
![]()
(2)柠檬酸异构生成异柠檬酸:在顺乌头酸酶的催化下,柠檬酸先脱水生成顺乌头酸,再加水异构成异柠檬酸。
![]()
(3)异柠檬酸氧化脱羧生成α-酮戊二酸:此反应在异柠檬酸脱氢酶作用下,异柠檬酸先脱氢再脱羧生成α-酮戊二酸。辅酶是NAD+,脱氢生成的NADH+H+经线粒体内膜上呼吸链传递生成水,氧化磷酸化生成ATP。这是三羧酸循环中第一次氧化脱羧。异柠檬酸脱氢酶是三羧酸循环的关键酶,是最主要的调节点,此反应不可逆。
![]()
(4)α-酮戊二酸氧化脱羧生成琥珀酰CoA:在α-酮戊二酸脱氢酶复合体催化下,α-酮戊二酸氧化脱羧生成琥珀酰CoA,脱下的2H由NAD+接受生成NADH+H+,氧化产生的能量一部分储存于琥珀酰CoA的高能硫酯键中,所以琥珀酰CoA为高能化合物。此反应不可逆,α-酮戊二酸脱氢酶复合体是关键酶。
![]()
(5)琥珀酰CoA转变为琥珀酸:琥珀酰CoA受琥珀酸硫激酶催化,将高能键转移给GDP生成GTP,自身转变成琥珀酸,这是三羧酸循环中唯一的底物水平磷酸化步骤。GTP又可将能量转移给ADP生成ATP。
![]()
(6)琥珀酸脱氢生成延胡索酸:在琥珀酸脱氢酶催化下,琥珀酸脱氢生成延胡索酸。FAD是琥珀酸脱氢酶的辅酶,接受脱下的2H生成FADH2。
![]()
(7)延胡索酸加水生成苹果酸:在延胡索酸酶催化下,延胡索酸加水生成苹果酸。
![]()
(8)苹果酸脱氢生成草酰乙酸:在苹果酸脱氢酶作用下,苹果酸脱氢生成草酰乙酸完成一次循环。NAD+是苹果酸脱氢酶的辅酶,接受氢生成NADH+H+。
![]()
三羧酸循环是乙酰CoA彻底氧化的途径。1分子乙酰CoA经三羧酸循环共两次脱羧,生成2分子CO2,这是体内CO2的主要来源;四次脱氢,生成3分子NADH+H+和1分子FADH2。NADH+H+和FADH2分子携带的氢经线粒体呼吸链氧化成水可产生9分子ATP;一次底物水平磷酸化,生成1分子ATP。故1分子乙酰CoA经三羧酸循环彻底氧化共生成10分子ATP。
三羧酸循环的反应过程如图7-4所示。
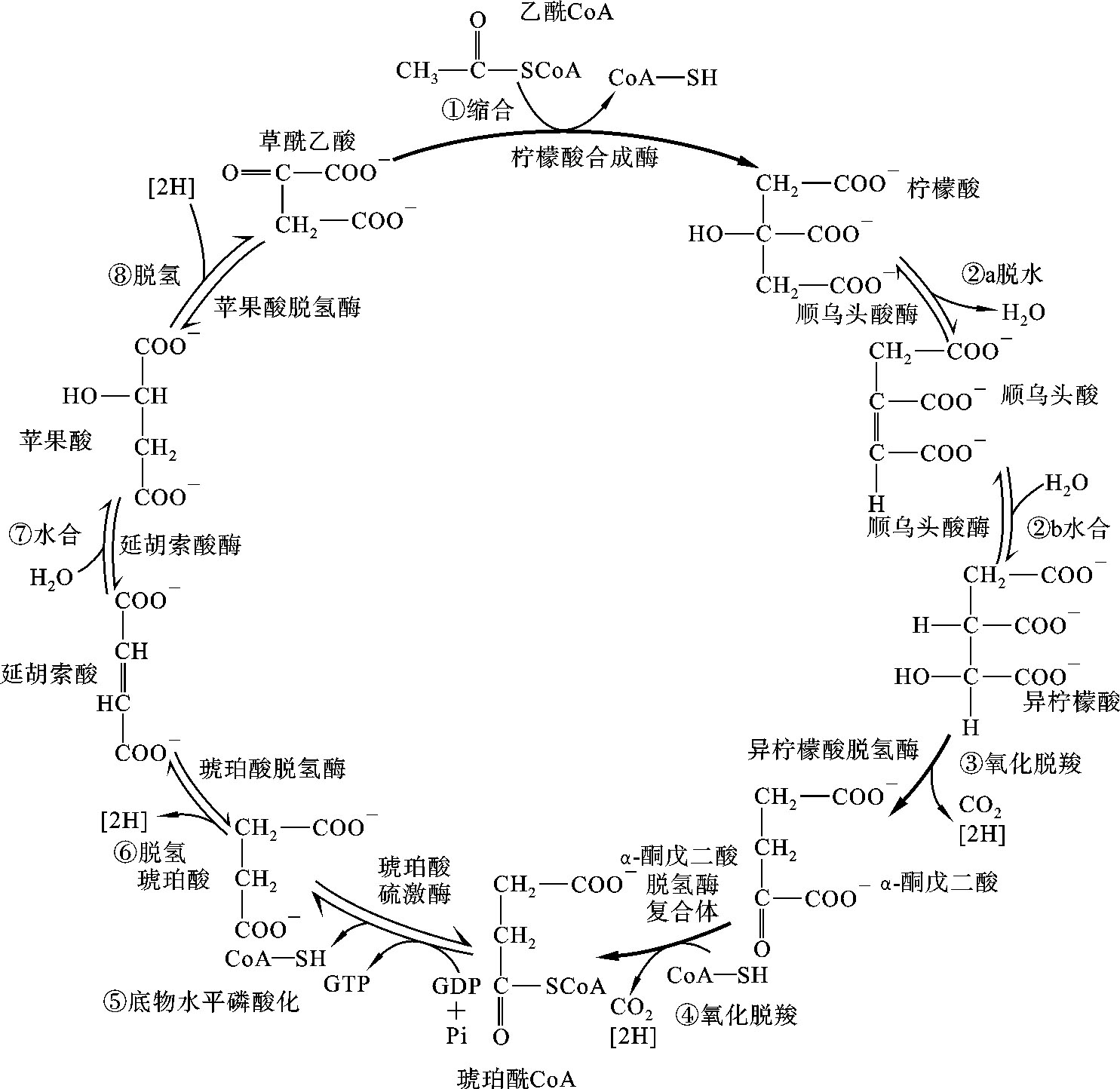
图7-4 三羧酸循环
(二)三羧酸循环的特点
(1)三羧酸循环是机体主要的产能途径:1分子乙酰CoA通过三羧酸循环经历了四次脱氢(三次脱氢生成NADH+H+,一次脱氢生成FADH2)、两次脱羧生成CO2、一次底物水平磷酸化,共生成10分子ATP。
(2)三羧酸循环中柠檬酸合成酶、异柠檬酸脱氢酶、α-酮戊二酸脱氢酶复合体是关键酶,催化的反应不可逆,故三羧酸循环为不可逆的单向循环。
(三)糖有氧氧化的生理意义
1.有氧氧化是机体供能的主要方式
1分子葡萄糖经有氧氧化生成CO2和H2O,能净生成30或32分子ATP(表7-1)。
表7-1 有氧氧化过程中ATP的生成
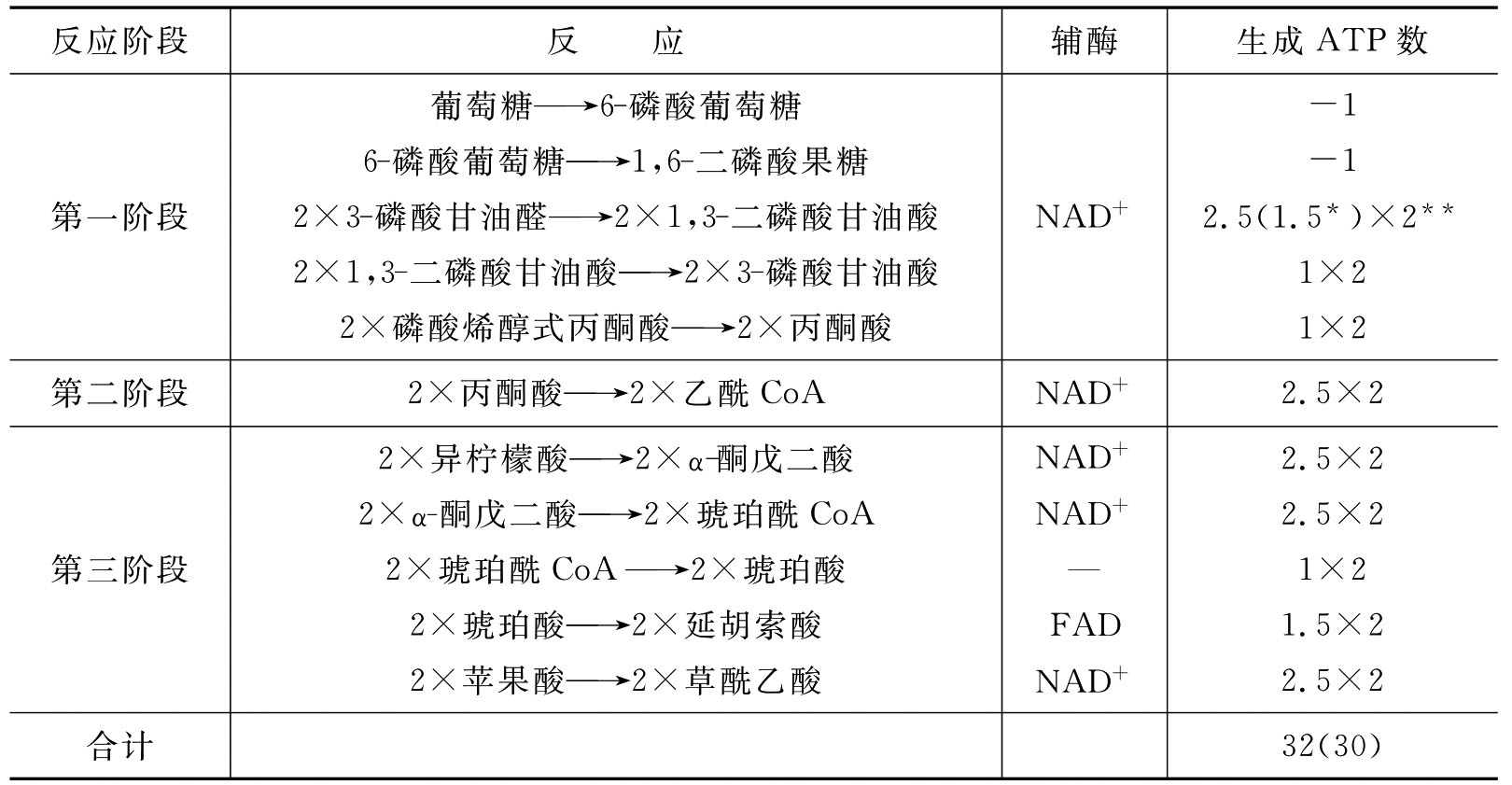
注:*根据NADH进入线粒体的方式不同,如果是α-磷酸甘油穿梭经呼吸链只产生1.5×2分子ATP;**1分子葡萄糖可裂解为2分子3-磷酸甘油醛,故×2。
2.三羧酸循环是体内糖、脂肪、蛋白质彻底氧化分解的共同途径
糖、脂肪、蛋白质经代谢后都能生成乙酰CoA,进入三羧酸循环彻底氧化,最终产物都是CO2、H2O和ATP。
3.三羧酸循环是糖、脂肪、蛋白质代谢联系的枢纽
三羧酸循环不是一个封闭的循环,而是一个开放的,与体内其他代谢途径相互联系、相互交汇的循环。如循环的中间产物α-酮戊二酸、丙酮酸及草酰乙酸通过氨基化作用生成谷氨酸、丙氨酸、天冬氨酸,而三种氨基酸又可经氨基酸代谢的脱氨基途径生成相应的α-酮戊二酸、丙酮酸及草酰乙酸,进入三羧酸循环。
糖代谢的中间产物乙酰CoA是合成脂肪酸的原料,氨基酸代谢的产物α-酮酸也可异生为糖等。
(四)糖有氧氧化与糖酵解的相互调节
巴斯德效应(Pastuer effect)是指在有氧的条件下糖有氧氧化能抑制糖酵解。这个效应是Pastuer在研究酵母菌葡萄糖发酵时发现的:在无氧的条件下,糖酵解产生的ATP的速度和数量远远大于有氧氧化,是产生ATP的主要方式,但在有氧的条件下,酵母菌的糖酵解作用受到抑制。这种现象同样出现在肌肉中:当肌肉组织在供氧充分的情况下,有氧氧化抑制糖酵解,能产生大量能量供肌肉组织活动所需,但在缺氧时,则以糖酵解为主。
在一些代谢旺盛的正常组织和肿瘤细胞中,即使在有氧的条件下,仍然以糖酵解为产生ATP的主要方式,这种现象称为反巴斯德效应。在具有反巴斯德效应的组织细胞中,其糖酵解酶系(己糖激酶、6-磷酸果糖激酶、丙酮酸激酶)活性较强,而线粒体中产生ATP的酶系活性较低,氧化磷酸化能力减弱,以糖酵解酶系产生能量为主。
三、磷酸戊糖途径
磷酸戊糖途径是葡萄糖氧化分解的另一条重要途径,它的功能不是产生ATP,而是产生细胞所需的具有重要生理作用的特殊物质,如NADPH和5-磷酸核糖。这条途径存在于肝脏、脂肪组织、甲状腺、肾上腺皮质、性腺、红细胞等组织中。代谢相关的酶存在于细胞质中。
(一)反应过程
磷酸戊糖途径在细胞质中进行。全过程分为两个阶段:第一阶段是氧化反应,产生NADPH及5-磷酸核糖;第二阶段是非氧化反应,是一系列基团的转移过程。
1.磷酸戊糖的生成
6-磷酸葡萄糖在6-磷酸葡萄糖脱氢酶及6-磷酸葡萄糖酸脱氢酶的催化作用下,经两次脱氢,生成2分子NADPH+H+,一次脱羧生成1分子CO2,自身则转变成5-磷酸核糖。6-磷酸葡萄糖脱氢酶是此途径的关键酶。
在这一阶段中产生了NADPH+H+和5-磷酸核糖这两个重要的代谢产物。
2.基团转移反应
第一阶段生成的5-磷酸核糖是合成核苷酸的原料,部分磷酸核糖通过一系列基团转移反应,转变成6-磷酸果糖和3-磷酸甘油醛。它们可转变为6-磷酸葡萄糖继续进行磷酸戊糖途径,也可以进入糖的有氧氧化或经糖酵解继续氧化分解。基本反应过程如图7-5所示。
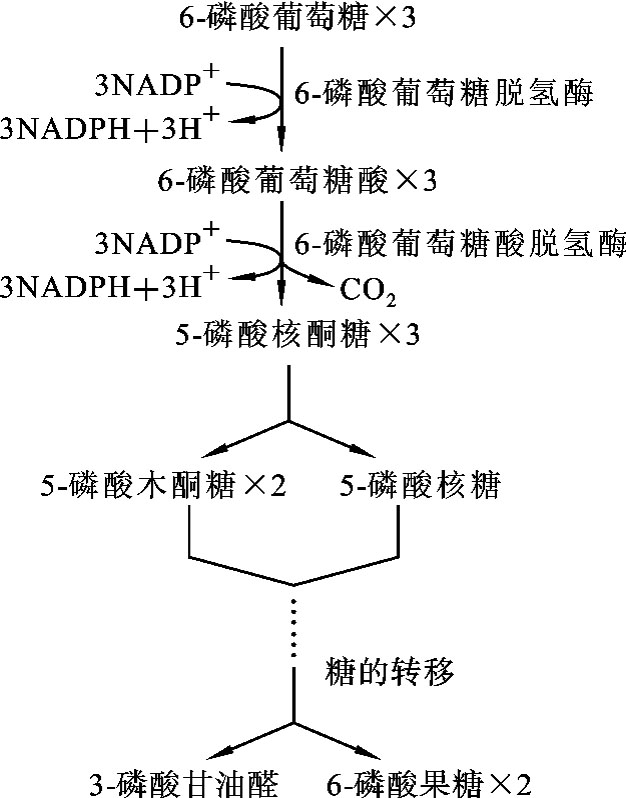
图7-5 磷酸戊糖途径
(二)生理意义
磷酸戊糖途径的生理意义不在于供能,它的主要生理功能是提供机体生物合成所需的一些原料。
1.提供5-磷酸核糖
此途径是葡萄糖在体内生成5-磷酸核糖的唯一途径。5-磷酸核糖是合成核苷酸的原料,核苷酸是核酸的基本组成单位。
2.提供NADPH+H+
NADPH+H+与NADH+H+不同,它携带的氢不是通过呼吸链氧化磷酸化生成ATP,而是参与许多代谢反应,发挥不同的作用。
(1)NADPH+H+作为供氢体参与脂肪酸、胆固醇和类固醇激素的生物合成。
(2)NADPH+H+是谷胱甘肽还原酶的辅酶:对维持还原型谷胱甘肽(GSH)的正常含量有很重要的作用。还原型谷胱甘肽是体内重要的抗氧化剂,能保护一些含巯基的蛋白质和酶类免受氧化剂的破坏。在红细胞中GSH能去除红细胞中的H2O2,以维护红细胞膜的完整性。H2O2在红细胞中的积聚,会加快血红蛋白氧化生成高铁血红蛋白的过程,降低红细胞的寿命;H2O2对脂类的氧化会导致红细胞膜的破坏,造成溶血。遗传性6-磷酸葡萄糖脱氢酶缺乏的患者,由于其磷酸戊糖途径不能正常进行,故造成NADPH+H+减少,不能使氧化型谷胱甘肽(GSSG)还原成GSH,造成GSH含量低下,则红细胞膜易于破裂而发生溶血性贫血。这类患者常在食用蚕豆后发病,故又称为蚕豆病。
(3)NADPH+H+参与肝脏生物转化反应:与激素、药物、毒物等的生物转化作用有关。
知识链接
蚕 豆 病
蚕豆、抗疟药、磺胺药等具有氧化作用,可使机体产生较多的H2O2。正常人由于6-磷酸葡萄糖脱氢酶活性正常,食用蚕豆或服用上述药物时,磷酸戊糖途径增强,可生成较多的NADPH+H+,导致GSH增加,这样可及时清除对红细胞有破坏作用的H2O2,而不会出现溶血。对于6-磷酸葡萄糖脱氢酶缺乏者,其磷酸戊糖途径不能正常进行,故NADPH+H+缺乏或不足,导致GSH生成减少。在正常情况下,由于机体产生的H2O2等物质不多,因此不会发病,与正常人无异。但当食用蚕豆或服用某些药物时,机体产生的H2O2增多,不能及时清除,从而破坏红细胞膜,而诱发溶血性贫血。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。









