



晋西中阳南川河中上游河沟植被调查简报(1955年10月)
序 言
1955年5月至8月参加了中国科学院黄河中游水土保持综合考察队的工作,对本调查区的植被作了初步的观察和记载。
南川河源于晋西中阳县刘家坪以西、老虎圪堆沟和弓阳镇以南王家山底附近,向北流下,至中阳县城会东川而继续北去,入三川河并转向西流后纳入黄河。调查区仅为中阳县城以南之中上游河段,到该县南之弓阳镇附近止,系一条南北路线长约30公里,东西宽约150米到200米的狭窄的河川沟谷,最宽处如车明峪一带300米,最窄处则在黄蒿岭及陈家湾以下约40—50米,河谷在中阳县城以北才逐渐宽广。整个调查区夹于中阳林区之间(图1)。
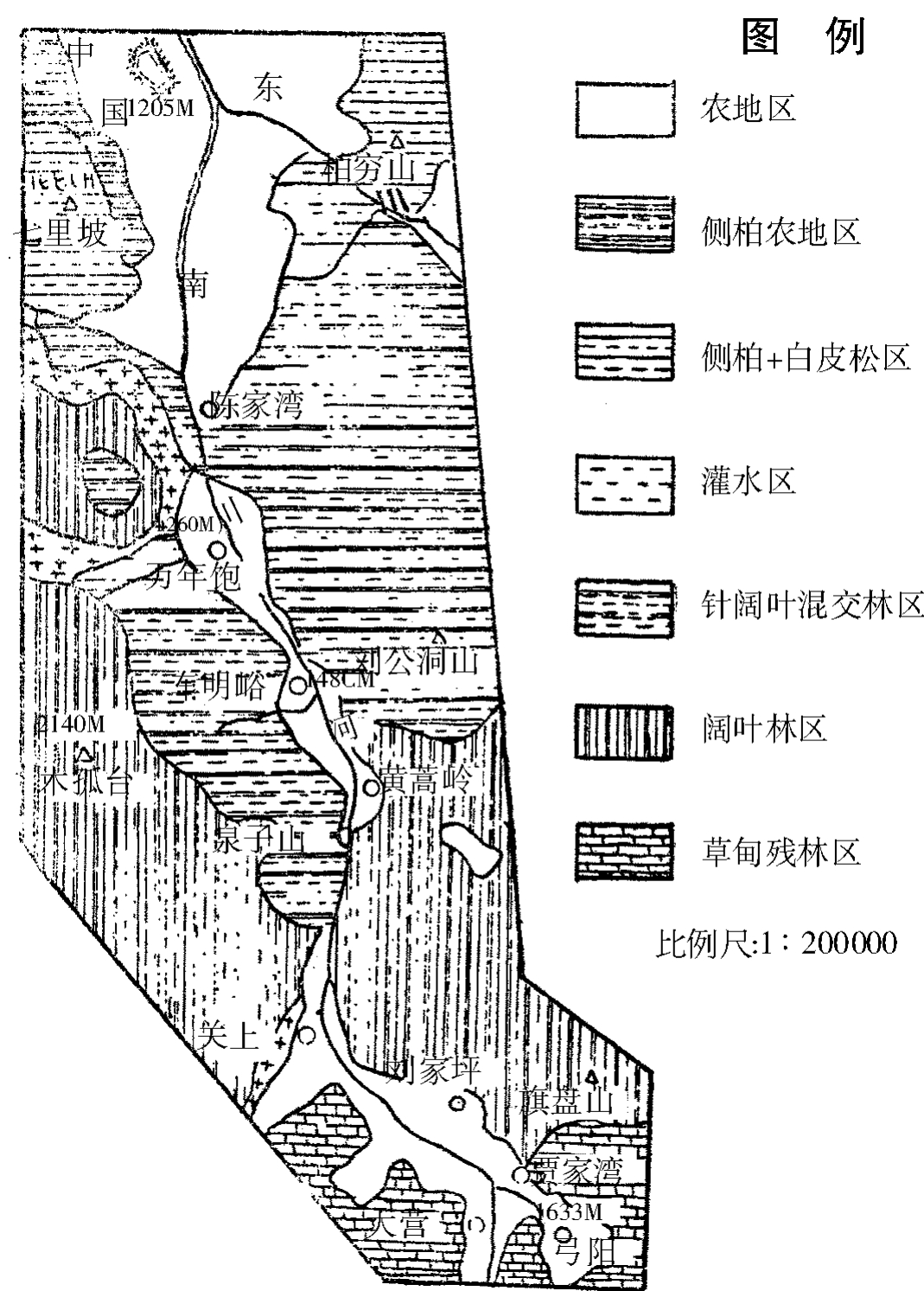
图1 南川河中上游河沟及其周围植被分布图
南川河中上游水小川窄,但又兼河川与沟谷之特点,因此河沟命名。
调查方式是适应整个工作部署而依次以中阳县、万年饱、车明峪、关上及弓阳镇等五个聚落所在的河段为据点,分别进行观察记载、样地调查、系列描写、口头访问及步度路线测量等工作。
一、环境综述
1.古老的侵蚀河谷地:地处陕北构造盆地的极东,呈单斜构造,乃吕梁北斜之西翼,山脉与基岩之走向基本一致,均为北北东—南南西,河沟斜跨其上(北西北—南东南),大都位于吕梁背斜的轴心部分,所露基岩均为各种古老的片麻岩、片岩和灰岩,整个河沟全在深切的露头之中。由于其拔海最低也在1200米以上,沟谷的相对高度也较大,短短的20公里,两侧笔直,雄伟壮观,其基本地貌之形成当在第四纪以前,此后的侵蚀和沉积仅排回其上,对原来已成面貌无大改变。
河沟海拔自1200米至1600米左右,但两岸依次有上述古老岩石露头的石质山峰,如七里坡、柏穷山、木弧台、刘公洞山、旗盘山及上山目等,一般高度在1600米以上,上顶山高2310米(图2)。
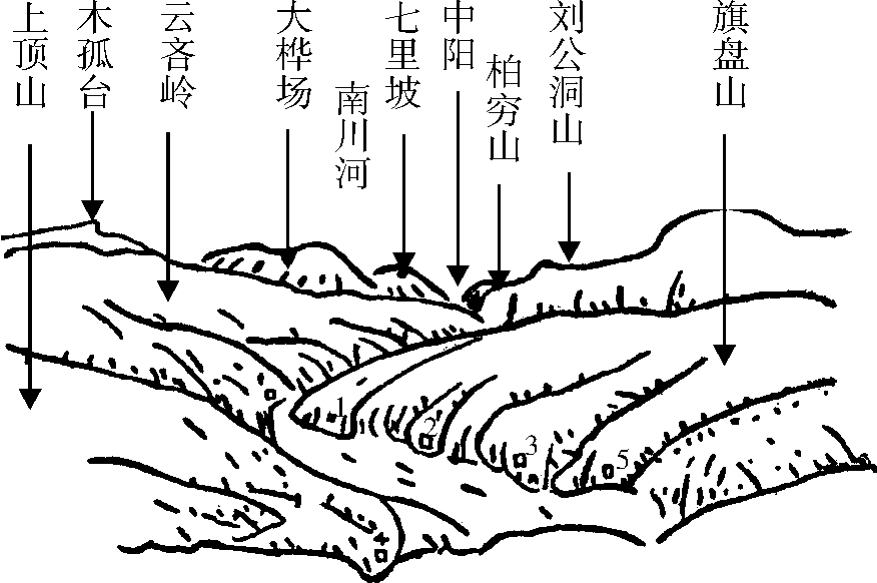
图2 弓阳跑马梁北望南川河谷
1.关上 2.刘家坪 3.神塘土焉 4.大营 5.贾家湾
河流自南向北,自源地之1655米到中阳县城之1205米间,高差450米,河流比降较大,但侵蚀极不明显,阶地也不显著,相反的河谷里的碎石沉积却不少。流域积水面积不大,水源仅来自岩隙,再加上流域内植被良好,“U”形谷中河曲蜿蜒。
河道平时没有支流,仅在雨季,车明峪沟偶有溪流汇入,在较大的支沟处有残存的冲积扇。
2.河沟纵向上的气候特点:中阳地处黄河中游,而黄河流域为典型的温带大陆气候,中阳气候受其直接控制,又由于森林植被覆盖,林区又较周围地区湿润,虽然范围不大而差异仍称鲜明,其特点是地势高,且南高北低,中上游为林区包围(图1),因此,虽长不及20公里而区别明显。
因缺乏气象记录资料,仅就访问及物候观察叙述如下:
河沟纵向上由北向南无霜期愈来愈短(由160天至125天),雪期则愈来愈长,冰冻也是北部较南部要早约30天,解冻则又相应推迟10天左右,同时地冻及冰冻深度也有明显差别。常向风为北西及北而又多以东及东南、南风降雨,雨多集中在夏秋,占全年雨水的25%—50%,最热及最冷时期也是依次递差。
这种气候反映在树木及作物生长也可以看出随高度由南向北的不同,最特殊的是油料作物荏子,只能种在中上游的中段,其南或北段均不适宜。
中阳降水1953年及1954两年平均为537.75毫米(因1954年雨水特多,因而多年平均则会更少些,估计450毫米左右)与北京(630.4毫米),太原(582.6毫米)及延安(521.1毫米),西安(557.2毫米)近似或稍逊。然而河沟调查区地处林区,因地高林密,林区降水则会因空气湿度较高而有所增多,如弓阳地区年雨量达600毫米左右。
河水温度就7月下旬记录也是近源低于远源,低于气温而更低于地温。
3.河沟横向上的土壤分布特点:由于河沟的上述自然特点,调查区基本上没有深厚而发育年龄较久的土壤,大都是近代冲积或坡积物发育而来的幼年性土壤。
纵向上,由弓阳到中阳除少数稍开扩部分的两侧及中阳以北有较厚的坡积性黄土外,所有的冲积土均甚薄,一般仅25—45厘米,最深者71厘米,然10—20厘米者不乏,其下部即为砾石层。无论坡积或冲积性土,其成土母质均为红色黄土,它们的共性是成土过程短暂,作用极不明显,碳酸盐反应还强,是一种持水保肥(矿质肥)能力较强,生产力较高的轻壤土,适于多种植物生长。
根据成土过程,母质来源及其自然条件,土壤在横向上的分布及其本身特点如下:
(1)碳酸盐生草潜育化土壤:分布在河床紧接河水的岸边,地下水位甚高处,厚度15—31.5厘米。土壤酸碱度7.0—7.2其上生长水边植物。
(2)河流冲积砂土:界于岸边与不明显的冲积阶地之间,带状或断带状随河身曲折而分布,一般厚度为30厘米左右。
据其发育历史及所处位置又可分成:多砾石的粗砂土、干河漫滩冲积砂土和湿河漫滩冲积砂土等三类,前者地干多砾石,基本不长植物,仅可见零星枯株;中者及后者均比较稳定的河漫滩上的冲积性幼年土,仅前者因地下水稍低土较干(表土因毛管现象并不干)等与后者的地下水位稍高而水分充足等有所不同。
(3)冲积土:分布于不明显的阶地上,南川河中上游分布较普遍。调查区因为土薄(40—70厘米)且干又远居民区而很少利用,下游已大多垦为农田。
(4)坡积及坠积性红色黄土所发育而来的土壤:原堆积于石崖顶部的红色黄土,后因各种原因而陆续坡积、坠积或风积于河沟近沟壁之两旁,厚度大,虽发育不久但内多碳酸盐和有机质(堆积或成土过程中卷入的)。稍加人工夷平即可利用(图3)。
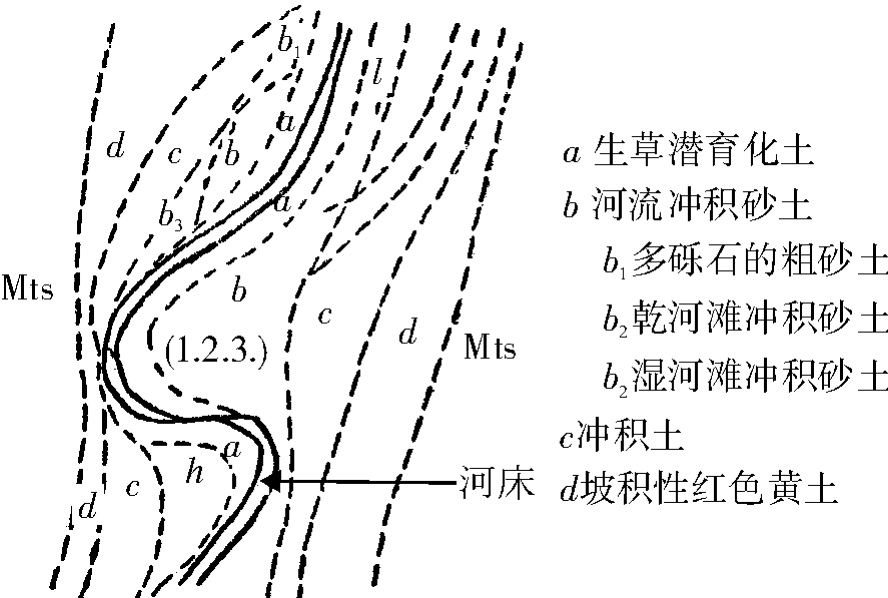
图3 河沟土壤分布示意图
此外在主支沟交会处的小型冲积扇上的冲积土,因每年都有一二次的冲积,肥力甚高,诸多利用。
4.从林区到耕作区的特殊的植被环境:以整个晋西植被分布而论,此处属吕梁山针阔叶林和灌丛草本植物区的低山灌丛针阔叶林复合群落亚区。这里称它为中阳林区。即北起中阳县城附近,南极县境东西以调查区为中心,向两边各达10公里左右,因此,河沟位于林区中心,除近河谷有少数被垦为农田外,中上游全为密林包围。由中阳索河源而向南依次分为三部分:
(1)中阳县城至陈家湾乃以侧柏和白皮松为主的针叶林复合群落;
(2)陈家湾至黄蒿岭乃以侧柏、白皮松、辽东栎、桦木和山杨、油松为主的针阔叶混交复合群落。
(3)黄蒿岭以南则为桦木、辽东栎和山杨为主的阔叶复合群落。
其间在陈家湾、石家沟、黄蒿岭和泉子山等处河沟甚窄,密林直逼沟壁。
这种植被环境对河沟其他自然条件有着直接控制作用,大凡河水流量少,流速少,变率平,无水土流失以及植物种子来源便利,因而易于繁衍等均同密林有关。
5.聚落分布及人文简况:南川河中上游本系林区周围诸地交通连系的孔道,加之森林发育,本来沿河沟一、二公里聚落存在,后来远在解放前受社会原因,居民大都外迁,然而至今尚有弓阳、关上、车明峪、万年饱及中阳等较大聚落,而且林区也有三五群居者。旧社会“逼上梁山”者大有人在;林源受到破坏,林木横遭摧残,林内有灾,林外必然受患。上游水土流失,下游沉积成灾,虽从中劳动人民也累积了不少斗争经验,然而国家财富之损失难以数计,这种情况解放后根本改变,林民生活有了保障,护、育、理、植林工作开展,为林区发展打开了广阔的前程,森林服务于人类的可能性才会最大发挥。
估计河沟居民400户左右,大约1500—2000人,合理利用林区,不仅林民生活完全可以保证,尚可支援国家社会主义建设对林业提出的部分需要。
综上所述,河沟水土流失基本停止,随海拔高度不同气候差异,土壤不同,利用方式也各端,植被的分布和演替当然也不同,随国民经济的发展其潜力也可适当发挥。
二、植被成分及其演替问题
1.植物成分
(1)横向上:以土壤因子为主导,土壤中又以土壤水分为首要,由河身向两岸分布:
Ⅰ.水中及水边植物成分:以蓼科、灯心草科、眼子菜科及菊科为主,调查期间有两棲蓼、酸膜叶蓼、萹蓄、泽泻、斑蓼、灯心草、眼子菜、大车前、水苦荬、旋复花、苍耳、百脉根、石龙芮、水门冬、薄荷、问荆、轮藻等31种。
Ⅱ.河漫滩植物成分(包括潜育土及砂土植物):
1)潜育土植物:包括水边植物的大部分,次外其中最多的是豆科、菊科、禾木科等科植物,调查期间有早熟禾、荩草、天兰苜蓿、齿草苜蓿、旋复花、野青茅、大车前、小龙胆及毛莨等18种。
2)砂土植物:在砾石块较多的砂土上菊科(如艾蒿、苍耳、铁杆蒿、牛尾蒿、茵陈蒿和阿尔泰紫菀等),豆科(如多花胡枝子、黄花苜蓿、扫帚苗等),禾本科(早熟禾一种、碱草、野青茅、狗尾草等),尤其是蒿属成分大量增多,调查期间所见23种中菊科占10种(其中蒿属6种),豆科占4种,禾本科达6种,在干河滩上大致情况同上,然豆科和禾本科植物数量相形增多,但种类则减少了,调查期间仅15种。
Ⅲ.冲积与坡积阶地植物成分:主要为已垦为农田的地区,成分将在下边田边及田间成分中记述。
(2)纵向上:表现上以温度为主导因子,而温度又受海拔高度及林区深度的影响,然本质则系因温度等多方面因子综合作用于生壤,从而导致其理化性质,微生物活动和土壤水分及肥力和差异的结果:
Ⅰ.自然植物成分:根据中阳、万年饱、车明峪附近及弓阳一带等四处分别记载的田边、田间和路旁植物看,不同的生态环境有着不同的植物种属,它们分别形成特殊的种属区系成分,虽然它们均系野生成分,但却与人类的生活活动有着极其密切的关系。
据上述四处不完全的记载,共得植物103种,分属33个科,其中最多为菊科(23种),其次为禾本科(13种),豆科(11种),蓼科(6种),再次为藜科、蔷薇科及唇形科(各4种),毛茛科、苋科、伞形科(各3种),其余均为一或二种。
虽调查时期有较大的差异(前后相差月余且其间旱涝和耕作情况有别),比较意义有一定的局限性,但仍可提出以下几点供诸者参考:
a.一般说各种蒿子到处可见,中、上游均有分布,田边、田间及路旁也随处可见。
b.小旋花、田旋花、苦荬菜、碱草等无论中、上游均为田间代表植物成分,而苋菜,地肤、黄花、沙蓬、虫实、蒺藜及柢枸杞等仅多见于中游,问荆、节节草、大车前等却见于万年饱或车明峪以南的上游田间。
c.碱草、艾蒿、叉明裸、茵陈蒿、车前及狗尾草为常见的田边植物成分、马蔺、问荆在万年饱以南才有,碱草、苦荬菜中游多见。
d.道旁杂草的代表如车前、蒲公英以及萹蓄、莳萝蒿、铺蔓委陵菜等,其中马蔺及蒲公英上游最多,蒺藜则中游多见。
e.此外还可以见到几种如飞廉,刺冲等几乎是调查区中游的优势杂草种类(至少是调查期间)。
Ⅱ.作物成分:以弓阳(上段),黄蒿岭(中段)及中阳(下段)三处的禾谷等作物品种调查作如下的比较:
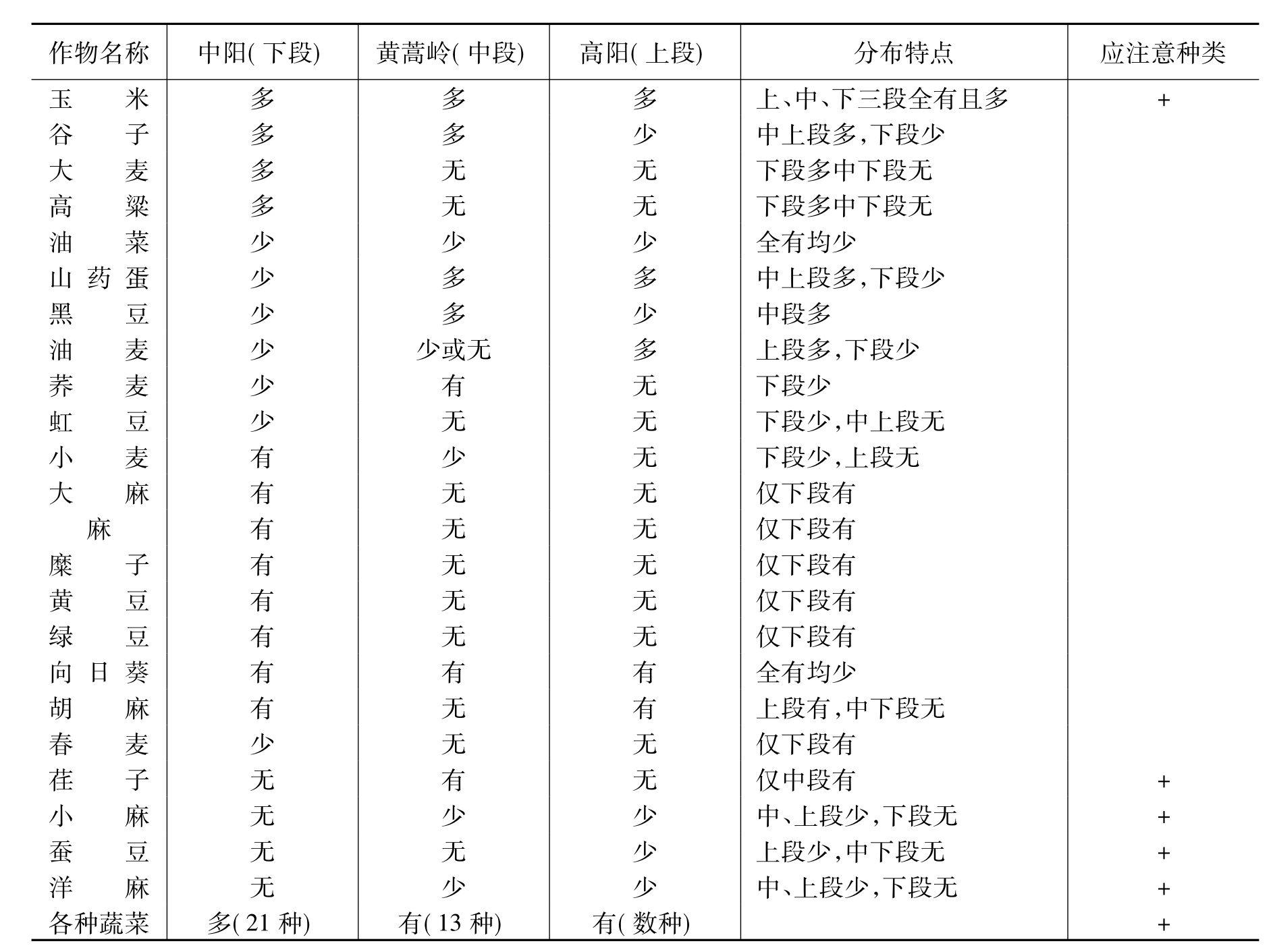
这里可以明显看出上、中、下三段作物的差异,如荏子只有中段有,油料及蔬菜作物则下段丰富的多,油麦上段多,中、下段倒少,只有玉米和向日葵是各段均有。
Ⅲ.其他杂草小灌木的分布特点:河滩上的植物分布也表现出上、中、下三段的区别:例如百里香只有关上以北才见到(可能为冲积土上的耐旱植物),水柏子黄蒿岭以南没有,只见于河沟之北段(温度的影响)、车明峪上下至万年饱生长最好。又如百脉根、高山绣线菊等亦多见于黄蒿岭以北。
2.植被描写
(1)水边杂草群丛组:分布在河边及河水中,河道非主流之一侧,形如绿色带子,调查期间的主要群落即为杂草群落,其成分30—35种,总盖度30%—40%。
万年饱河滩样的为例:优势种为早熟禾一种,灯心草一种(土岸)、鬼针草(土边水中)、荩草(土岸),此外还有泽泻、两蓼、眼子菜(均水中)等共33种,生活一般中等,总盖度30%(图4a.b),其植被的生物学特性是多年生种子繁植,株小,根不发达,保护组织不发达,易萎蔫等。
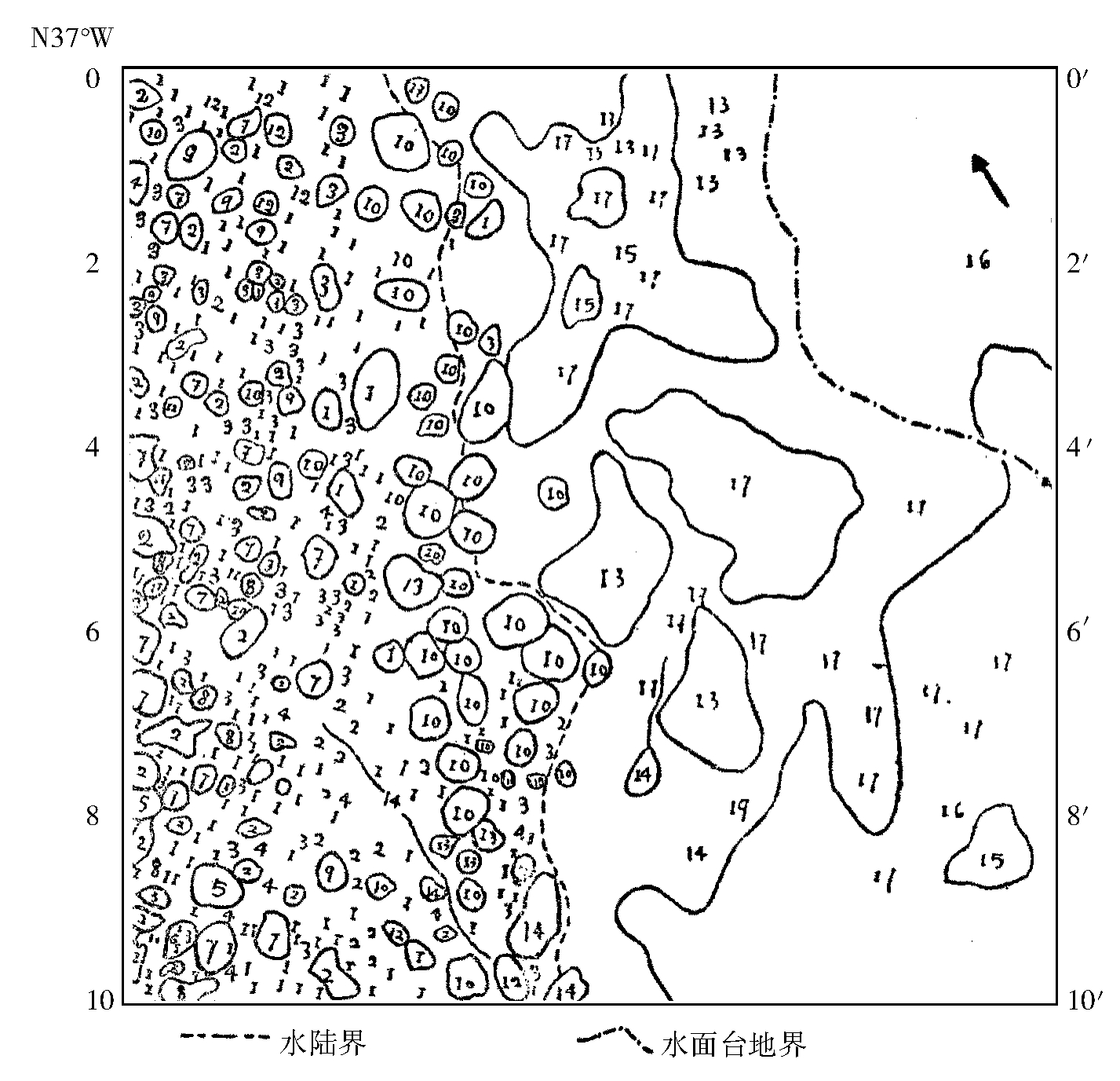
图4.a 万年饱水边杂草群落样地投影图
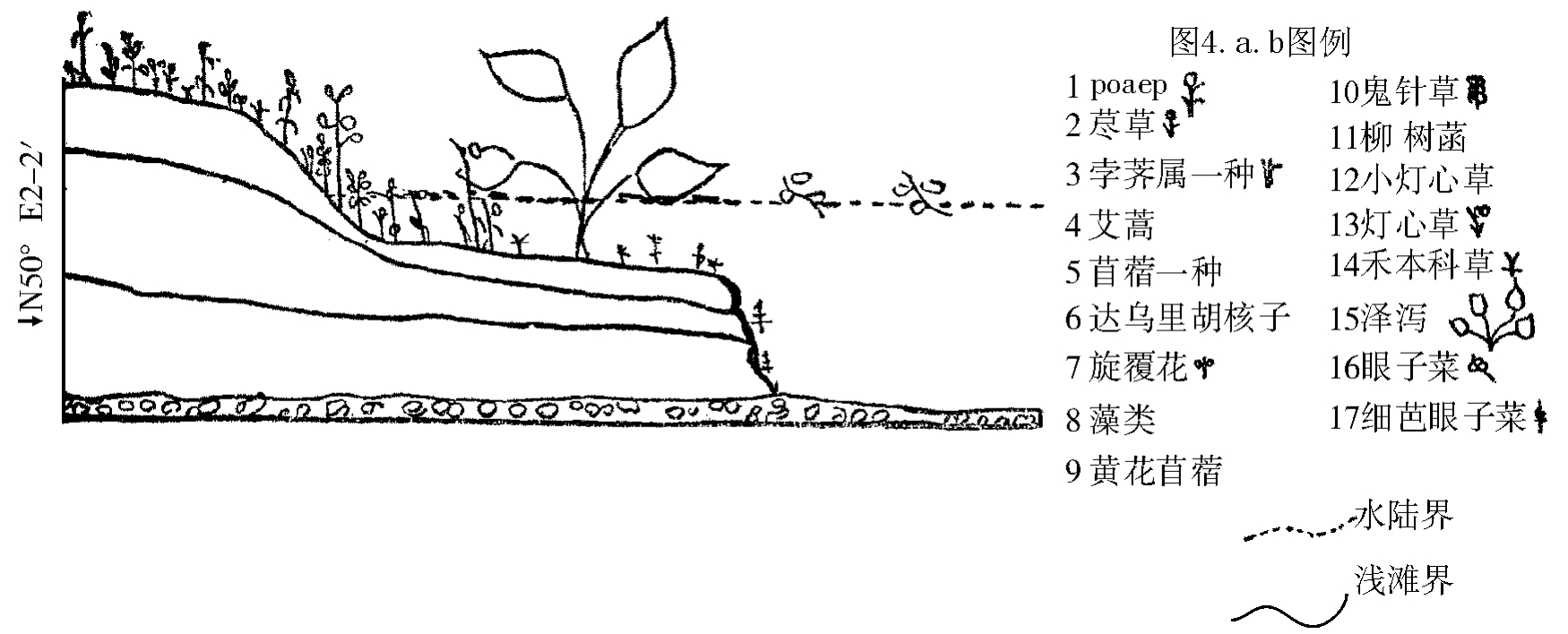
图4.b 万年饱水边杂草群落系列图
(2)河滩杂草群丛组:随河滩历史、土层、水分等条件而具有各种不同群落:
Ⅰ.杂草群落:河间滩地上,表土沙质,牲畜践踏较重地上分布。特点是种数少,优势种不明显,大都一年生,偶有柳、榆树苗。
万年饱村下所见者总盖度65%,共有26种。优势成分有早熟禾一种,齿草苜蓿、旋复花、荩草及艾蒿、铺蔓萎陵菜等。
Ⅱ.苔草一种群丛:分布并不多,万年饱以北岩石削壁下,乃轻微流动泉水边的沮洳地的碳酸盐中度生草灰化土的植物群落,成分简单,植株矮小,但生长茂密,总盖度75%以上,组成仅17种,全为水生及湿生喜耐阴湿植物。其中苔草一种最占优势,总盖度67%以上,生活力极强,此外还有问荆、荩草、盘龙参及水柏子等。
Ⅲ.铁秆蒿+野菊群丛:涨水线以上的河漫滩上,地瘠土薄(10厘米),卵石裸露,土多砂质,虽有植物达30种,但盖度不大。
车明峪调查是:成分32种,盖度仅12%(调查时水淹,可能偏低),以成丛的铁秆蒿和株小数多的野菊占优势。
Ⅳ.禾本科一种群丛:湿润的近水边而非水边处,形如绿色带子,种数少而生长尚密茂。
刘家坪南贾湾的样地是:成分19种,总盖度90%,优势种(禾本科一种)盖度17%,但其多为Cop’,此外还有野青茅、车前、齿草苜蓿等。调查时正值花期及果期(7月22日),外貌尚称美丽。
Ⅴ.碱草+鹅冠草+野燕麦群落:土层较厚(1米以上),干燥坚实的冲积阶地上,以禾本科草占优势。
关上、泉子山一带所见者总盖度40%,组成26种,除群落名称的优势种外,尚有毛茛及其他禾本科植物6种、菊科8种、豆科3种。
该群落所在系十多年的放荒地,今后当随禾本科草的减少而为灌丛所演替。
Ⅵ.碱草+艾蒿群落或称旱河滩草本群落:土层虽厚但因牲畜践踏而表土坚实,水分蒸发,其上仅长旱生植物。
贾家湾所见群落,调查期间外貌一片黄褐色,点缀以灰绿色草堆。成分15种,总盖度约为20%,均高20厘米以下,只有碱草可达50厘米。群落中有零星酸刺出现,乃其过渡象征。
Ⅶ.百里香+艾蒿群丛:亦为土厚坚实的干旱阶地上的植物群落。关上北五里的上会村所见其种类稀少(16种),盖度仅25%,但从其中的酸刺、黄刺玫等出现及其生活力但盛看,其演替趋势将为灌木所代替,似乎无疑。
总之,河滩地的草木群落的形态结构及动态是极其复杂的,以各种途径和不同速度向灌丛及乔木群落过渡。
(3)河滩灌木群丛组
Ⅰ.黄刺玫—铁秆蒿+茭蒿群丛:分布在土壤经过一定发育的河流阶地上,乃草木群落向灌丛演替过渡类型。
泉子山以北因地处深林、种子来源方便和环境郁闭的原因,这种群落发育良好,其中有不少林下植物,如高山绣线菊、柴氏子、多花胡枝子、大龙胆、小叶唐松草等。虽种类较复杂达47种但盖度仅40%;其中灌木14%、草本25%,可能系人为不良影响的结果。
Ⅱ.绢柳+酸刺—杂草群丛:水丰土厚的环境里,湿生及中生植物混生,除绢柳及酸刺外尚有旋复花、苍耳、盘龙参、地榆、黄芩、石竹等组成灌木杂草群丛。
Ⅲ.酸刺群丛:多分布河曲凸岸河滩阶地上。上会村所见者总盖度50%—90%,成分29—64种,以酸刺占绝对优势,株高3—4米,生活力很强,约5—10年的树龄,树杆具刺,密集,人牲不入,草本优势为禾本科(7种)、菊科(5—16种)、豆科(4—5种),群落中有散生乔木,如青杨(盖度1%—2%),绢柳及榆树等小苗。
(4)散生乔木及乔木群丛组
Ⅰ.散生乔木成分:整个河沟散生着榆树、柳树、小叶杨等,此外还偶尔可见辽东栎和油松的植株。
Ⅱ.柳树—酸刺群丛:环境较湿的阶地便有由酸刺群丛演变而来的乔灌木过渡型群丛,柳树深根易在60厘米下取得养料(表土贫瘠)。车明峪所见者盖度可达95%,成分27种:其中乔木层盖度30%、柳树占25%(50平方米中有13株);灌木层盖度15%,酸刺为优势;草木盖度达80%,优势种不明显。柳树有日趋繁荣之势。
Ⅲ.小叶杨—乌柳群丛:河岸边碳酸盐弱度潜育化土壤,土层薄而水分充足,群丛有人工抚育现象,但林相极不整齐。车明峪的群丛总盖度95%,成分38种:其中乔木层生活力强,但稀(25平方米内有小叶杨4株),灌木层密茂,盖度70%,除乌柳(占60%)外还偶有茶条槭等;草木层异常稀疏,盖度5%—6%,且无明显的优势种。
Ⅳ.小叶杨群丛:由人工抚育而来(砍去酸刺),多在农田及聚落近旁,黄蒿岭有抚育6年的该类群丛,成分50种、盖度仅45%:其中乔木层占30%,除小叶杨外还有柳树;灌木层主要如酸刺平均高2.5米、盖度16%;草木层则以铁秆蒿、艾蒿为主,盖度约15%。
(5)其间动态演替关系之探索:复杂而多途,如图所示:
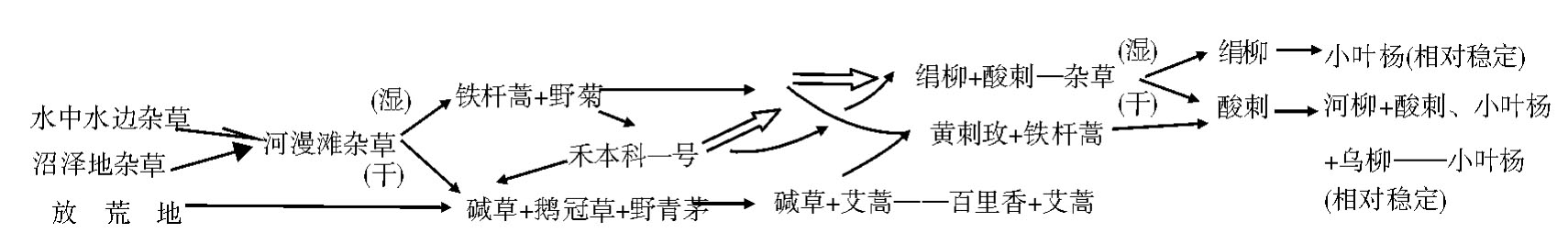
晋西中阳南川河中上游河沟地带植物群密演替系列示意图(所列植物名均为群丛或群落名称)
至于小叶杨群丛之进一步发展,据推测则可能因为所在中阳林区的位置而定,如继续不受人为影响,则此后的动态节律与整个林区植被一致。然这次调查中未作进一步探讨。
河沟自然植被演替基本与中阳林区相似,即由草木而灌木、乔木,不过因河沟水分及热力分布的差别而所经历的具体道路不同。观察过程中所见到分布和演替上有如下几点:
水边及水中植物在水分充足的情况下普遍分布且比较稳定,其演替随河床的变更而变更,这种变更有两种形式:其一为河流起主导作用的洪水冲积改变原状者,这种情况则需时较暂,如贾家湾滩地便因王家山底之洪水而发生变化;另一则为环境相对稳定,长期发育中由植被引起环境变化,进而导致植被变化者,其需时较久而短期内变化不显,植物居变化之主动地位。其变化趋势一般是向远河床之群落类型方面发展。
潜育土植物可由草本占据极长时期,在环境改变过程中,在种子来源不同的情况下,酸刺等灌木可以侵入,引起代替,经灌木阶段而乔木阶段,然而同时乔木亦可直接侵入,通过比较短暂的时间,由乔灌木混生而长成河沟杨柳乔木群落,如加人工抚育,则乔木成长更快。
冲积阶地上虽表土比较干燥,但因直接接近河水及地下水面,若土层稍厚即可发育起草本、半灌木(蒿)、灌木(酸刺)等不同的混生,而后成长木本群落。
可利用的积冲地在放荒后并不经过长期的草本阶段而直接由蒿类(首先是茵陈蒿),在三、五年后即过渡到酸刺灌丛。
此外,人类活动对植被演替也发生重大影响,河沟不少杨柳群丛均为人工抚育而成便是证明。
三、几种植物个体生态或小群落的描写
河沟植被中除上述“彼此惯于一起生长的,具有一定种类成分和一定群落外貌的植物种的总体”(阿略兴语)的植物群落外,值得注意的还有一些植物的个体生态及“由繁殖特点”或“由植物群落中无植被的地面”或“植物环境的条件”所决定的,个别的植被片断的小群落。由于工作关系未能全面细致地观察,仅就所见,列举几个:
1.眼子菜的生态观察:生长在比较停滞的淡水中,除根部着生在泥土中以外,整个植株顺河水流向而分布,既不靠河底又不浮水面,流速过急的水中不能生长,过浅的滩中也少分布,因而多在流速不大的河曲外主流之一边的岸边,万年饱村下河床边较多分布,弓阳北贾家湾的河沟中原为水流处,后因洪水而河床变更,原来生长眼子菜的河水边改变为沼泽性质的,因而眼子菜的生活力不强,植株短小。
2.水苦荬的生态观察:水苦荬为玄参科多年生草本,常见于水中及水边,茎中空,叶对生,一般多见于滞水中。但有时在流水中亦可见到,这时其根之大部在水中,只有约1/3的须根下部在水中的泥里(图5),此乃其对流水冲刷的不良条件的适应本能,根入土方向与流水相反。根之弯曲可能系洪水期作用之结果。
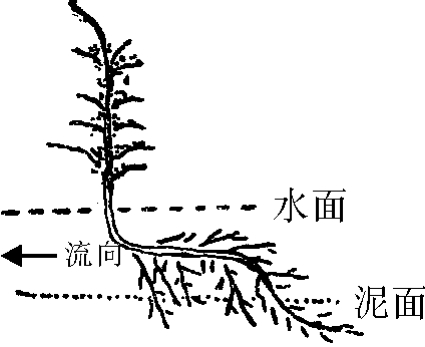
图5 水苦荬生态示意图
3.草小群落的描写:成丛密生在地下水位较高的河岸边,镶嵌成带状或条片状,其所在为冲积的粗砂砾石经过强度或中度潜育化的土壤上。覆盖度50%左右,伴生植物有水蓼、野青茅、鬼针草、荩草、水门冬、大灯芯草、水苦荬、泽泻、旋复花、大车前等杂草,种类不少,但个体数目不多,有些还是偶见种植物。凡与其伴生者多生长不良而发育较快,如野青茅15—20厘米高即开花,旋复花8厘米就开花等。
根以极发达的地下茎相连,地下茎节间长短不等,随小环境而不同从1—6厘米,节间有鳞片,抱茎而生,须根在生枝叶的地方密生着,长约5—6甚至10厘米,且许多须根相互交错成为毡织状的一团,密生在5—7厘米深的沙土上,其地下茎(或叫“主根”)所在处土壤大都强度潜育化,其他植物确难进入,这是伴生种植株少或生长不良的主要原因。
愈离水边愈近,则其生长甚密,向水边或水里愈远,则分布愈疏,一般急流中没有,流水中仅有稀疏分布,多在河岸水边20—100厘米之内分布,25—40厘米之间,生长甚健旺。
4.鬼针草简记:生在水边,特别是常水位与涨水位线间最多,伴生种很多,均水生及水边植物。根的形态结构简单,共有3—7条粗而光滑的支根,根毛及须根极少,根约与地上部分等长,多成群生长。
其所在常为潜育化的砂土,常同其他植物群生,如在松软的土上植株不易直立。
5.荩草小群落描写:荩草为禾本科小草,生长在湿河滩上,近水而且平坦,土层多黏质、较厚而没有砾石,只在河水边约1米的平滩上长着,也大致呈带状分布,在薰草之内侧,分布幅度不等,从20到100厘米,偶有更宽者,一般盖度可达70%左右(与其他伴生种一起),生长很密,植株小而数目极多,其分布见图6。
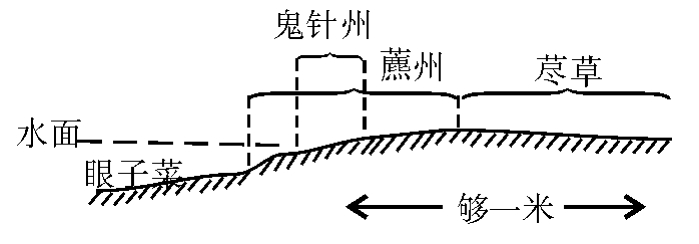
图6 眼子菜、鬼针草、草及荩草之分布示意图
伴生植物有大车前、早熟禾一种,禾本科一种、旋复花,天兰苜蓿、石龙芮、薄荷、苍耳及齿草苜蓿等。
值得注意的是荩草在这里生长慢而且很小,7月中仅有2厘米,8月初也不过6厘米,但又生长甚密,可能与气候有关。
6.百里香小群落描写:稍干而空旷的河滩上,由于其走茎繁殖而成堆生长,盖度可达30%以上,一般为直径25—40厘米的团块状,断续连接而成大约可达6—10平方米的大片分布。其伴生种有碱草、茵陈蒿、背铺萎陵菜、达乌里胡枝子、米口袋、阿尔泰紫菀、紫花地丁、达乌里龙胆、铺蔓萎陵菜及隐子草等旱生和中生植物。
百里香植株走茎而附地生长,株高5.5—6.0厘米间,叶小对生。花亦小粉红色(稍带玫瑰色)、唇形花冠、四数、上位花、子房下位。花期5月底至8月初,种子繁殖但就其生长状况看似以营养繁殖为主。根为直根,亦可为不定根,当随所在土壤的薄厚而定:如土层极薄为1—15厘米以内,则直根退化,不定根发育;如土层在35厘米左右,便可发育起15—20厘米内直根了。
小群落的外貌为细密的黄绿色小草,上缀以小的玫瑰色总状花序,极其美观。
在酸刺群落中亦可看到百里香小群落,因环境不同(稍荫),其伴随植物亦有所区别,如细叶远志、铁秆蒿、碱草、萎陵菜、米口袋等。总之大致与灌丛外貌相同唯多了几种中生植物,似乎说明酸刺群落可能是这种小群落的插入种进一步发育而来的灌木群落。
7.水柏子的生态:灌木,叶上生白色小盐点,生水边,肉质。所在环境土厚约20—30厘米,其下为砾石层,砂壤及粗砂壤。生长在河滩上,地上部分忍受流水冲击,地下部分受砾石层大小石块的挤压(大者直径有达15厘米)。伴生种较杂,大凡水边、河滩植物都有。
其根系为多年生的大规模的“地下茎”(实则原为地上部),主根深入到砾石层下(地面下40厘米深处还有,然土层仅厚21厘米)夹于碎石粗砂间,支根极少。地下茎遍布在地面下20厘米的土层中,纵横分枝,异常紊乱,但主根与地下茎的关系是沿流水的流向一侧弯曲,所见竟有顺延达6米者,此间牛羊可食。
8.铺蔓萎陵菜的个体生态:路旁植物或盐渍化的河滩植物,常见于表土坚硬的地方。
其主要特点是匍匐茎生长及其营养繁殖,有时长可达55—65厘米,叶绿、茎红、花黄,形成特异的外貌,每段匍匐的节间长8—12厘米之间,每节上23个小的掌状深裂的叶子,其下就着地而生不定根,但也并非每节都生根,因为它们相互交错而并非每一节都可着地。分布地其他植物不易进入,因而伴生植物寥寥,仅在其外围土质松软或水分充足处始有苍耳、旋复花、齿草苜蓿、灯心草等。
尾 声
晋西中阳南川河中上游河沟地带系一古老的侵蚀河谷,乃吕梁背斜的西翼,拔海在1200—1600米间,周围受石质山峰环抱,既受温带大陆东岸季风气候的控制,又受周围中阳林区气候的显著影响,由于地势南高北低,气候亦依次差异。土壤由于沉积类型及具体发育条件(水分)等的不同,河谷横断面上有着明显的差异,由河床向两岸土层由薄变厚,水分由多量变成中量。河谷系交通要道,人文影响明显。
河沟植被与其特定之自然环境形成有机的统一,无论种类成分和群落类型既受整个中阳林区之作用,又与河沟条件一致。其突出表现在成分上的纵向差别和群落上的横向殊异,野生植物如此,栽培作物亦然。
从其自然植被与栽培作物品种及其动态演替结合晋西山区尤其中阳地区居民的经济生活来看,其利用之潜力仍很优异。大凡林木之适量供应,绿化树种之采集,作物菜蔬药材之经营,山区林业副业之开发利用等均有一定之实际意义。由于其自然条件较特殊在研究河沟植被恢复演替上也有一定的理论价值。
1955年9月20日—1955年11月20日北京大学初稿,1956年11月西北大学学术讨论会报告,原载1957年《西北大学学报》。
附录:本文所列之植物名录(某些作物学名未列入)
轮藻科(CHARACEAE)
轮藻一种(Chara sp.)
木贼科(EQUISSETACEAE)
节节草(Equisetum ramosissimum Desf.)
问荆(E.arvense L.)
松科(PINACEAE)
白皮松(Pinus bungeana Zucc.et Endl.)
油松(P.tabulaeformis Carr.)
柏科(CUPRESSACEAE)
侧柏(Platycladus orientalis CL.)France
麻黄科(EPHEDRACEAE)
麻黄(Ephedra sinica Stapf.)
杨柳tu(SALICACEAE)
青杨(杨树)(Populus cathayana.)
山杨(P.davidiana Dode.)
小叶杨(P.Simonii Carr.)
乌柳(Salix cheilophila Schaeid)
旱柳(河柳)(S.matsudana Koidz)
杞柳(S.purpurea L.)
绢柳(S.viminalis L.)
桦木科(BETULACEAE)
桦木(白桦)(Betula Pendula)
山毛榉科(FAGACEAE)
辽东栎(Quercus liaotungensis koidz.)
榆科(ULMAEAE)
榆树(Ulmus pumila L.)
蓼科(POLYGONACEAE)
雨棲蓼(Polygonum anphibium L.)
萹蓄(P.aviculare L.)
酸膜叶蓼(P.lapathifolium L.)
斑蓼(P.nodosun Pers.)
藜科(CHENOPODIACEAE)
沙蓬(沙米)(Agriophyllum anenarium Bieb.)
虫实(绵蓬)(Corispermum declinatum Steph.ex Stev.)
地肤(Kochia scoparia Schrad.)
沙蓬(猪毛菜)(Salsola collina pall.)
叉明科一种(S.sp)
苋科(AMARNIACEAE)
苋菜(Amaranthus mangostanus L.)
石竹科(CARXOPHYLLACEAE)
石竹(Dianthus chinesis L.)
毛茛科(RANUNCULACEAE)
毛茛(Ranunculus japonicus Thunb.)
石龙芮(R.secleratus L.)
小叶唐松草(Thalictrum elegans)
蔷薇科(ROSACEAE)
紫氏栒子木(灰栒子)(Cotoneaster Zabelli Schaeid.)
委陵菜(Potentilla chinensis Ser.)
朝天委陵菜(P.supina L.)
黄刺玫(Rosa xanthina Lindl.)
地榆(Sanguisorba officinalis L.)
高山绣线菊(Spiraea alpina pall.)
三裂绣线菊(S.trilobata L.)
豆科(LEGUMINOSAE)
米口袋(Gueldenstaedtia verna(Georgi)Boss.)
达乌里胡枝子(Lespedeza dahurica(Laxm.)Schindl.)
多花胡枝子(L.floribunda Bge.)
百脉根(Lotus corniculatus L.)
天兰苜蓿(Medicago lupulina L.)
齿草苜蓿(黄花苜蓿)(M.falcata L.)
蒺藜科(ZYGOPHXLLACEAE)
蒺藜(Tribulus terestris L.)
远志科(POLYGALCEAE)
细叶远志(Polygala tenuifolia Willa.)
槭树科(ACERACEAE)
茶条槭(Acer ginnola Maxim.)
柽柳科(TAMARICEAE)
水柏子(Myricaria dahurica Ehrenb.)
堇菜科(VIOLACEAE)
紫花地丁(Viola mandshurica W.Becker)
胡颓子科(ELAEGANCEAE)
酸刺(Hippophae rhamnoides L.)
龙胆科(GENTIANACEAE)
小龙胆(达乌里龙胆)(Gentiana squarrosa Lebeb.)
大龙胆(秦艽)(G.macrophylla pall.)
旋花科(CONVOLVULACEAE)
小花旋(Calystegia hederacea Choisy.)
田花旋(Convulvulus chinesis Ker-Gawl.)
紫草科(BOPBAGINACEAE)
刺仲(Lappula echinata Gilib.)
唇形科(LABIABIATAE)
薄荷(Mentha haplocalyx Brig.)
黄芩(Scutellayia baicalensis Georgi.)
百里香(Thymus mongolicus Roan.)
荏子(Perilla frutescens(L.)Britton.)
茄科(SOLANACEAE)
枸杞(Lycium chinenso Mill.)
玄参科(SCROPHULARIACEAE)
水苦 (Veronicaanegalis-agvaticaL.)
(Veronicaanegalis-agvaticaL.)
车前科(PLANTAGINACEAE)
车前(Plantago asiatica Linn.)
大车前(P.major L.)
菊科(COMPOSITAE)
莳萝蒿(Artemisia anethoides,Veber.)
茵陈蒿(A.capillaris Thunb,)
茭蒿(A.giraldii pamp.)
铁杆蒿(A.sacrorum Ledeb.)
牛尾蒿(A.dubia Wall.Ex Bess.)
艾蒿(A.argyi Levl.et Vant)
阿尔泰紫菀(Aster altaicus Willd.)
鬼针叶(Bidens parviflora Willd.)
飞廉(Carduu crispus L.)
野菊(Chrysanthemum indicum L.)
旋复花(Inula britanica L.)
苦 菜(Txerischinensis(Thunb.)Nakai.)
菜(Txerischinensis(Thunb.)Nakai.)
蒲公英(Taraxacum mongolicurn Hand-Mazz.)
苍耳(Xanthium Japonicum Wills.)
眼子菜科(POTAMOGETONACEAE)
眼子菜(Potamogeton octandrus Poir.)
窄叶眼子菜(P.pectinatus L.)
芝菜科(JUNCAGINACEAE)
水麦冬(Triglochin palustre L.)
泽泻科(ALISMATACEAE)
泽泻(Alisma orientale L.)
禾本科(GRAMINEAE)
鹅冠草(Agropyron chinensis Ohwi.)
碱草(Aneurolepidium chinensis Kit.)
荩草(Arthraxon hispidus(Thunb.))
油麦(Avena sativa L.)
野青茅(Deyeuxia clarion arundinacea(L.)Beauv)
隐子草(Cleistogenes sp.)
早熟禾一种(Poa sp.)
狗尾草(Setaria viridis(L.)Beauv.)
高粱(Sorghum vulgare pers.)
小麦(Triticum aestivum L.)
玉米(Zea mays L.)
莎草科(CYPERACEAE)
 草(Scirpus triqueter L.)
草(Scirpus triqueter L.)
灯心草科(JUNCACEAE)
灯心草(Juncus effcusus L.var.decipiens Buchen.)
小灯心草(J.sp.)
百合科(LILIACEAE)
黄花(Hemerocallis minor Mill.)
鸢尾科(IRIDACEAE)
马蔺(Iris enssta Thunb.)
兰科(ORCHIDACEAE)
盤龙参(绶草)(Spiranthes arstoteria Merr.)
参考文献(文中已注明者未列入)
[1]B·H·苏卡且夫等著、李继侗译:地植物学研究简明指南,科学出版社,1955年。
[2]科学院土壤研究所编辑:土壤调查手册,科学出版社,1955年。
[3]崔友文编著:华北经济植物志要,科学出版社,1953年。
[4]胡先骕著:经济植物手册(上、一二、下一),科学出版社,1955年、1956年、1957年。
[5]B·B·阿略兴著:Л·B·库德里亚绍夫改编、傅子桢、王燕合译:植物地理学(上册),高教出版社,1954年。
[6]科学院西北农生所编:陕西太白山种子植物名录(专利)。
[7]中阳县人委所编:有关资料(内部油印资料)。
[8]谢寅堂:西安野生植物调查报告,西北大学学报,1957年第1期,197—213页。
[9]北京大学生物系:1955年,晋西生产实习队,晋西中阳植被调查报告(未刊)。
[10]人民革命军事委员会气象局:中国气象资料,1951年。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











