



1.2.1 研究内容
1.2.1.1 岩石圈、水圈、大气圈作用于生物圈及其造成的影响
岩石圈、水圈、大气圈作用于生物圈,对生物圈造成的影响,又可通过生物圈的响应(如古生物地理学与古生态学的反演)而识别这些影响,并应用于地球系统科学。
1.生物地层学(Biostratigraphy)详见第2章。
2.生态地层学(Ecostratigraphy)详见第3章。
3.古生物地理学(Paleobiogeography)和古生物-气候研究
(1)古生物地理学。详见第5章。
(2)古生物-气候研究。大气环境控制生态系及其演化,因而古生物适应古气候,并可反演古气候。由于生物-气候反馈机制的研究,已可将生物及衍生物用于古气候研究。例如,陆地植被的反照率、有机碳的埋藏(生物矿物是大气CO2的一个汇)、影响云团形成的二甲基硫化物等生物化合物的产生、海洋生物产量和Fe和P供应之间的关系等,是全球变化的重要参数;孢粉、造礁珊瑚、赤道冰川和地下水的同位素和化学数据,是证明在末次冰盛期时热带变冷的第一手证据;地史时期各典型气候带的生物群,为古气候分带及其变迁提供了证据(赵锡文等,1992)。目前,由古生物反演古气候的研究通常归入古生物地理学或古气候学中,而未独立成古生物气候学。
1.2.1.2 生物圈作用于岩石圈、水圈、大气圈及其造成的影响
1.生物成岩作用(Biodiagenesis) 详见第6章。
2.生物成矿作用(Biometallogenesis) 详见第7章。
3.生物地球化学 研究生物地质作用过程中各种化学元素的性状以及生物迁移、富集、沉淀化学元素的能力和方式。当前着重两方面的内容,即元素的全球循环和生物圈的影响。生物圈质量虽小,但物质和能量的循环速度极大,如某些细菌在适宜条件下繁殖,只经36h就能把地球表面盖上一层,所以生物地球化学作用是地球表层系统中最强的化学作用。生物新陈代谢,吸取元素又释放元素,从而使地壳元素迁移和重新分配,它控制土壤和沉积物中大多数的Fe,S,P,C,N等元素(和同位素)的分布;海洋生物新陈代谢过程对海水中其他主要元素(如Ca和Si)和次要元素(如As,Mo和 U)亦具有明显的影响;生物活体代谢或死亡后形成的有机质,尤其是有机流体(含有机质的气体或液体),由于其胶体性、还原性、络合性、吸附性等,对元素的活化、迁移、再分布有重大作用。生物地球化学循环如生源要素(C,N,P)的循环,以及生物、有机质、有机流体在矿物分解(有机产物可使硅酸盐矿物的分解速率增加2~100余倍)、矿物沉淀、土壤形成和沉积物成岩成矿等过程中所起的作用,无疑对全球变化产生关键作用,对于了解现代和古代地球环境变化过程十分重要。
1.2.1.3 分子地球生物学
分子地球生物学(Molecular geobiology)是NRC(2000)中创用的词,意指分子生物学、分子古生物学(Molecular paleontology)和地学相结合的研究。本书将其作为一个独立的内容,是考虑到:①它的研究对象特殊,是生物和古生物分子(生物标志化合物、生物大分子、基因),甚至包括前生命的有机质,这就大大扩展了生物地质学的研究时间和空间;亦因此,其研究方法也与前不同;②分子地球生物学正在蓬勃发展,成为地球系统科学的重要组成部分,分子生物学是生命科学的前沿和当前主流,我们可以预期分子古生物学亦将成为古生物学的前沿和主流,应单独列出;③生物地质学并不完全包含分子地球生物学,如后者中分子生物学与流体地学的交叉即不属于生物地质学。由于②和③两点,分子地球生物学将来有独立出去的可能。分子地球生物学研究内容包括∶
1.生物出现前的化学生物学和生命的起源 包括生物大分子的起源、
细胞膜的起源、遗传密码的起源、磷的作用、粘土表面的有机反应等,现已认识到生物出现前的生物大分子在38亿年前即已形成。例如,硫酯和乙酸等分子在早期地球演化的地球化学环境下即已生成,核苷酸和氨基酸也已合成,这为生命起源提供了可能的线索。
2.从地质体追索生命的演化、特别是早期演化 沉积岩中生物标志化合物的同位素组成特征,现已可用高精度仪器(如GC-C/TC-IRMS)加以测定和描述。这样就可能追溯生命演化中重要新陈代谢过程出现的时间、顺序及其作用。分子证据表明,早在25亿年前,蓝藻就在生产大量氧气;至少在27亿年前,具有复杂结构的真核细胞生物就可能已经出现,这比已知的最古老的真核化石早5亿~6亿年。
3.演化树、分子钟 大多数动物的核心进化基因是相同的,植物亦是如此。根据生物分类单位的基因相似性,可建立起包含全部生物门类的分子演化树。由于相当一部分蛋白质分子进化速度具有恒定性,可以根据生物DNA之间的差异,对分子演化树的各分支标定时间,建立起分子钟。演化树、分子钟与年代地质学相结合,并与化石记录中新类别的出现年代相检验,可建立起包含演化树各主要分支事件的地质年代表。从最早的多细胞动物出现到脊椎动物各纲的分化,都可以一一标识。地质和化石数据可用来校准分子钟;分子钟也可以帮助建立其他缺乏丰富化石记录的系统谱系。
4.分子地层学和全球变化的分子标志 作为地质历史时期生物有机体遗留下来的有机分子,特别是其中的分子化石,能精细记录各种不同时间分辨率的全球气候和植被变化信息。研究载体已涉及全球变化的诸多关键载体,如海相沉积物、湖泊和泥炭沉积物、冰芯、黄土-古土壤系列、洞穴石笋、树木年轮等。例如海洋沉积物长链不饱和酮的不饱和度与海水表面温度的定量关系,全球性古火灾(如白垩纪、古近纪)的分子标志等。利用分子化石这些特征,可以在传统的哑地层(如红土)中划分对比地层,用气候事件及灾难事件的分子标志对比地层等,从而形成了分子地层学。
5.极端环境中的生命过程 在极端环境条件下(如“深部生物圈”)发现的古细菌类(嗜盐菌、嗜热酸菌、极端嗜热菌),已经构成生物界的新的大类,大大拓宽了生命多样性的概念;在南极冰盖几千米深处、沿洋中脊的活火山口中和pH值为零的流体中也已经发现了微生物。板块俯冲带中可能存在地球上最深的、由化学氧化还原反应释放的能量(而不是太阳辐射能)维持其生存的生态系。同时,对极端环境中的现代生命过程的认识,不仅在生物技术应用上有巨大潜力,而且对于进一步理解生命起源和地外生命都有重要的理论意义。这些极端环境生命过程的研究很多都是在分子水平上进行的。
6.环境污染的分子标志 在当代环境污染研究中,已明确提出了环境污染的化石生物标志物和人类活动标志物。前者指化石燃料和沉积物中的分子化石造成的现代环境污染,典型的例子如海洋中的石油泄漏造成的污染。后者指人类在利用这些分子化石过程中所引起的环境污染,如人工合成的DDT,PCB,PAH等的有机污染。这些有机污染物已影响到了大气圈、水圈(冰冻圈)、生物圈和土壤圈,有关这些污染源的形成过程和消除都离不开对分子化石本身地球化学性质的研究。
上面1~4属于分子古生物学和分子地层学(Molecular stratigraphy),详见第4章。
1.2.1.4 生物圈与地球层圈相互作用的规律性研究
1.生物演化中的环境因素问题 生物演化的理论大体主要有三类。它们对环境在生物演化中所起的作用观点不同,因此,生物演化中的环境因素成了一个问题。现简述三类的观点如下:
(1)进化的综合学说(Evolutionary synthesis)。又称新达尔文主义(Neo-Darwinism)。它与达尔文传统的进化论一脉相承,特别强调自然选择的作用,认为自然选择决定了演化的方向和速度,并在群体遗传学和生物统计学的基础上重新解释了自然选择。这是上个世纪生物演化的主流理论。自然选择来自对环境的适应,故此说重视环境的作用,但侧重环境和生物的渐变。
(2)间断平衡论(Punctuated equilibria)和新灾变论(Neocatastrophism)。间断平衡论为Eldredge与Gould(1972)所提出,它与传统的演化论主要区别有二。其一,传统的演化论强调演化是渐进过程,在时间(纵)-性状演变(横)坐标上呈斜线的形式,如图1.2b。间断平衡论则认为演化是突变(间断)与渐变(平衡)的结合,强调大多数物种是在很短的、地质上可忽略不计的时间内,通过分出新枝系而完成的。这个迅速的分枝过程叫成种作用(speciation),在上述坐标上表现为接近水平的横线,即在时间值几乎不变的条件下,性状有了突然的变化。物种形成后性状基本稳定,只有缓慢的渐进变异,叫种系渐变(pheletic gradual evolution),在上述坐标上表现为接近于垂直的纵线。整个演化呈折线状,如图1.2a。传统演化论认为演化量(性状变异总量)是渐变积累的总和(图1.2c中的1~5),即种系渐变是演化的主流。间断平衡论则认为主要的演化量由成种作用构成(图1.2c中的两段横线),它才是演化的主流。
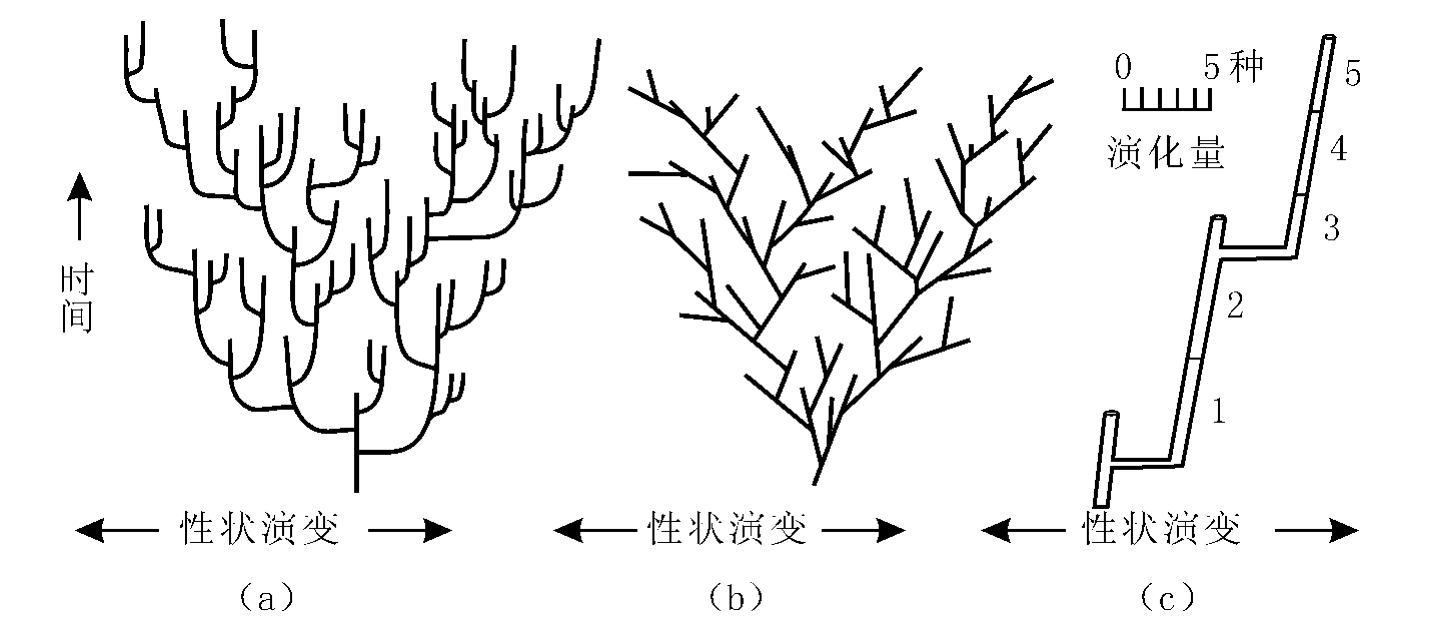
图1.2 生物的两种演化模式(据Stanley,1979)
(a)间断平衡论模式(b)渐变的模式(c)突变和渐变造成的演化量比较
其二,传统演化论强调外因(环境)是动力,外因具体表现为自然选择,它使生物适应变异,并决定演化的方向和速度。间断平衡论则着眼于内因(基因和染色体的变异),对于外因的着重点亦有转移(强调地理隔离)。它认为形成新种的原料是基因突变,突变是随机的,不一定沿着更好的适应的方向进行,而是无定向的,只要对适应无害就可能遗传。它又强调由父种地理分布区边缘上被隔离的孤立小居群中形成新种。因为在隔离小居群中产生的突变不致像在大居群中那样,因基因交流而失去特征。其中大多数虽被淘汰,少数仍能被保留下来,形成新种。
由于20世纪多次发现了在很短时间内突然辐射繁盛的生物群(如澄江动物群、热河生物群等),以及其后的6次重大灭绝事件(见下文),使多数古生物学家相信,演化并非稳定地逐渐趋向完善,间断是生命史上多次发生的事实,从而接受了间断平衡论。
新灾变论为Schindewolf(1954)所提出,被现代灾变学说引用而加以发展。它与居维叶所提出的灾变论的区别,一是强调地外(宇宙)因素,二是完全抛弃了居维叶的神创论。灾变不同于突变。突变主要指演化的加速(成种作用),即外界因素不中断演化过程,只是诱发和加大了进化速度和烈度。灾变则是演化的中断,即外来因素突然打断了演化进程,往往表现为在一个很短时间内发生多门类生物的灭绝。
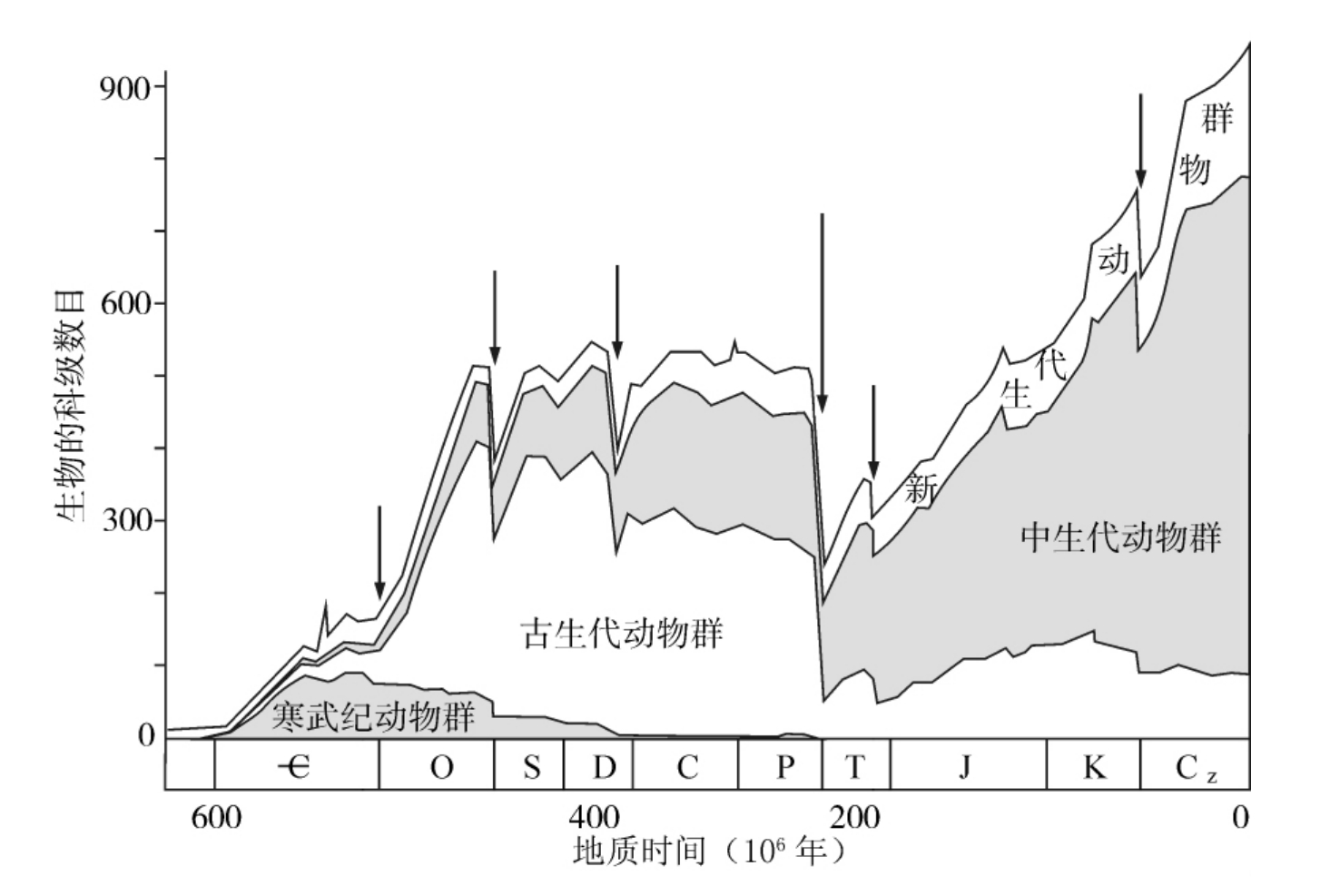
图1.3 地质时期生物科级数目变化反映的6次(箭头所指处)重大灭绝事件(据Sepkoski,1984)
显生宙以来的6亿年中共发生过6次大灭绝,这6次的时间为寒武纪末、奥陶纪末、泥盆纪末、二叠纪末、三叠纪末和白垩纪末(图1.3),此外还有一些规模较小的灭绝。长期稳定与短期灭绝相交替,生物演化突然被中断,这用渐变论无法解释。特别是Alvarez等(1980)报道了白垩纪末生物灭绝处Ir的异常高含量,认为这次灭绝是地外星体撞击地球的结果,因为Ir在地壳内的背景值很低(小于0.1×10-9),只在地外天体中富存(为地壳的104倍)。他们的发现引起了新灾变论的复兴。新灾变论提出,生物大灭绝是由于宇宙和地球演化中出现过一系列剧烈的灾变所致,它们具有短时、高能、突发的特点,如超新星爆发、太阳耀斑、彗星撞击、火山爆发、缺氧事件、地磁极反转等。有地外因素与地内因素之分。白垩纪末的“星体撞击说”,因为在地层中找到灾变事件留下的一些记录(如痕量元素Ir,Os等的异常,稳定同位素异常,微球粒,冲击石英,撞击坑等),现已得到较多的认同,作为新灾变论的实证。
间断平衡论和新灾变论都承认环境的作用,但特别强调生物和环境的突变(由事物本身的渐变—突变规律造成)和灾变(由事物以外不能预见因素造成)。
(3)分子进化中性学说(Neutralism)。此说为木村资生(M Kimura,1968)所创。他主张:在生物的分子水平上,进化不是由自然选择作用于有利突变所造成,而是在连续突变压力下,由(对自然选择)呈中性或接近中性的突变的随机固定所造成。中性学说承认自然选择在表型进化中起作用,但否认它在分子进化中起作用,认为决定生物大分子(蛋白质、核酸)进化的主要因素是突变压和机会,从而把自然选择看成是进化的次要因素,也就基本上否认了环境的作用。
由上可见,三种主要进化学说对环境因素的观点不同,它们可能是各自摸到了大象的一部分,我们在生物地质学研究中,需要对之进行处理和验证。
2.生物与地球的协同演化(co-evolution of organisms and earth)
宏演化基本模式(basic model ofmacroevolution):古生物突变或灾变式演化的新思潮,彻底改变了以渐变论为基调的演化基本模式。生物演化不仅是由简单到复杂,由低级到高级的前进式发展,而且是由大量灭绝、适应辐射、稳定发展相交替的阶段式发展。由此,在20世纪90年代,引发出显生宙生物大爆发、大灭绝、大复苏、大辐射的宏演化模式。生物的爆发、灭绝、复苏和辐射成为生物宏演化进程中有机联系的四个重要过程,它们构成地质历史和生物演化的重大转折时期。
在生物演化规律的研究中,人们发现在生物宏演化中无论渐变、突变或灾变,都是与它们生存环境的稳定、变化或剧变相一致。生物界的每一次突变都与地球环境的重大变化密切相关。例如,晚元古代多细胞生物群的繁盛,是在地球从冰冻的雪球期解冻之后发生的;寒武纪初的生命大爆发,其先决条件之一是当时大气圈具备了足够的氧气;奥陶纪志留纪之交的生物灭绝与奥陶纪末冰川时期后的迅速转暖有关;古、中生代之交剧烈的全球变化与生物大灭绝同时发生;中、新生代之交恐龙的灭绝,则可能与外星体撞击造成的环境剧变有关。
另外,在当代全球变化的研究中,人们亦发现生物圈演化与地球各圈层密切相关。生命的起源与演化受地球环境,特别是气圈、水圈和岩石圈演化的控制,而生命过程又反过来影响固体地球表层环境及大气圈和水圈的变化。现代大气圈(氧气占22%)主要是由海洋浮游生物和陆地植被的新陈代谢活动造成;土壤主要由微生物作用于风化产物造成。地球之所以不同于其他行星,就是因为它有生物圈。因此,生命过程与地球环境的演化是相互作用、相互制约的协同演化过程。当代地学已进入地球系统科学研究阶段,物理、化学与生物过程是地球系统中圈层相互作用的三大基本过程,所以生物圈与其他圈层的相互作用,即生物与地球的协同演化,是地球系统科学的基本内容之一。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











