



“噪声”的词根源自拉丁文“恶心”(nausea),这直率地表达了我们有时候对噪声的那种情绪反应。它是令人烦恼的、不可预料的事件,干扰了事物的自然状态,驱使我们离开和谐,也的确引起我们恶心的内态感。生物本身的细胞和分子组成就是噪声性质的,正因为此,生物机体是一种不完美的机器就不足为奇了。但是我们的生活能够没有噪声吗?噪声是不是一种反常的基本特征,使得我们在朝向完美的过程中失去常规,但同时也使我们有别于无生命的机器?当我们的探讨对象是活生生的脑时,这个基本的问题就很值得讨论了[1]。
“状态”一词具有广泛的含义。神经元的上状态和下状态是作为慢于1 Hz慢振荡的伴随而被发现的,对慢振荡的研究其实起源于对觉醒-睡眠周期的探索。与上状态相关联的有神经元“高电导状态”,即皮层神经元接受强烈突触冲击,处于更加去极化的高电导状态。在整体水平则有脑功能的“激活”状态。甚至把麻醉剂作用下所产生的活跃及静止时期的交替,称为“上状态”和“下状态”。所以,我们看到了从神经系统噪声直到脑功能状态之间的连续体(continum),它们之间可能存在着某种基本性质上的相通之处,这是非常值得介绍和研究的。
离子通道和突触的活动一定伴随有噪声和可变性,因而是脑功能无法逃避的元素[1]。这里有两个未获解决的最主要问题:①在何等程度上,噪声扭曲和限制了信号的传输?②在一个不可预测的世界里,噪声和可变性如何提供了关键而基本的丰富信息,使得易化及应付复杂的适应性行为成为可能?本章拟介绍近来的实验证据,说明功能性神经网络的活动如何伴随着膜电位及锋电位的时机而强烈波动,追踪噪声和可变性的来源及其后果,讨论无噪声的神经元信号通信,以及在大规模功能性神经网络中噪声的有害及有利方面。在某些情况下,噪声的可变性与行为可变性是并肩而来的,并且由此增加了行为的丰富多彩及可适应性。相关证据将可能开启今后研究之新途径[1]。
噪声是一种随机的信号扰乱,它造成信息加工的基本问题,影响神经系统功能的所有方面。然而神经系统内噪声的性质、数量和影响,最近才以定量的方式被提了出来。实验和计算机方法显示,多噪声来源促使细胞和行为测试的逐次(trial-to-trial)变异。本章从分子到行为水平回顾神经系统里面噪声的来源,并显示噪声如何导致逐次变异,强调噪声如何影响神经元网络,以及神经系统如何对抗噪声有害效应的原理,并简要讨论噪声的潜在好处[2]。
虽然以往的评论都聚焦于一般的神经元易变性,但我们这里聚焦于直接联系到噪声的一些话题。噪声渗透到神经系统的各个水平,从感觉信号的知觉开始,直到产生运动反应,而且对信息加工提出了基本问题。近年来一些问题得到了研究,如噪声在多大程度上存在,以及噪声如何形成神经系统的结构与功能。在本章中,我们首先考虑中枢神经系统噪声的性质、数量及其对中枢神经系统的效应。因为脑的目的是接受和加工信息,同时发出动作以应答信息(图8-1),所以我们考虑噪声对行为回路各个水平可变性的贡献,考察噪声如何影响运动行为。最后,我们讨论神经系统在知觉、抉择及运动行为当中运用什么样的策略来对抗、补偿或引起噪声。由于所涉及的系统及水平很多,我们不能够提供非常完整的评述。相反,我们要挑出一些反映更普遍情况的特例,说明噪声给中枢神经系统带来的约束及局限性,也讨论噪声的好处[2]。
图8-1 神经系统的行为回路和各阶段所存在噪声的总结(彩图见图版此处)
(a)感觉噪声的来源包括信号转导,这里由光感受器及其信号放大级联来代表。(b)细胞噪声的来源包括兴奋性膜、突触传递和网络相互作用中的离子通道。(c)运动噪声的来源包括运动神经元和肌肉。在所显示的行为作业(抓一个皮球)中,神经系统必须在有感受、信息加工和运动的噪声存在情况下动作。(图引自[2])
外部感觉刺激本身具有内源性噪声的性质,它们可能是热力学或量子力学性质的。例如,所有形式的化学感受(包括嗅觉和味觉)都受热力学噪声的影响,因为分子通过弥散过程以随机速率来到受体,而受体蛋白质准确计数信号分子的能力是有限的。类似于此,在视觉方面,光子被光感受器所吸收,其速率是由泊松过程决定的。这使得视觉的对比敏感度有其物理上的极限,当光线弱的时候这个极限减小,因为在光照明程度低的时候,仅有少数光子抵达光感受器[2]。
在知觉的第一站(图8-1a),感觉刺激中所含的能量(通过光的吸收或气味物质与配基的结合)或者机械信号(例如听觉当中毛细胞的运动)转化为化学信号。往后的转导过程把感觉信号放大,直接或间接地通过第二信使级联反应,把感觉信号转变成为电信号。任何感觉噪声不论是已经存在着的,还是在放大过程中产生的(转导噪声),都可以逐次增加可变性。因此,噪声水平为后续的信息加工规定了知觉阈值,因为经过放大之后,比噪声弱的信息不再能与噪声相分辨。这一点受到数据处理不等同性(data-processing inequality)理论的强有力支撑,此理论认为,对往后阶段的加工(即使它们是无噪声的)比之前阶段所提供的,不能够提取更多的信息。因此为了减少噪声,机体经常要在加工的第一阶段即感觉阶段付出很大的代谢与结构代价。例如,苍蝇的光感受器需要消耗10%的静息代谢,而它用在眼视觉的消耗占飞行负荷的20%[2]。
由于加工信息的细胞机器之随机过程,每个神经元的噪声会累积起来。噪声还可由于非线性计算及神经网络相互作用的结果而进一步增加。在神经元中,从生物化学和生物物理学水平来看,有许多随机过程在那里起作用,包括蛋白质的产生和降解、离子通道的开放和关闭、突触小泡的融合,以及信号分子弥散而与受体结合。通常人们无保留地假定,这种大量随机元素的平均可以有效地去除随机性。然而,这个假定需要重新评估。神经元实现包括高增益放大及正反馈的高度非线性操控,因此小的生物化学和电化学波动(当在分子水平上考虑系统的时候,我们应用“波动”这个词,其意义与噪声是可以互换的)能够明显改变整个细胞的反应。例如,当膜电位接近放电阈值的时候,动作电位的产生变得对噪声高度敏感。粗大的神经元结构例如乌贼巨轴突(其直径可粗达1 mm),被广泛地用来研究神经机制。这些结构的这种尺度似乎在功能上具有决定性意义,由于大量信号分子的参与,随机波动的确就被平均掉了。然而许多神经元是纤小的,小脑平行纤维的平均直径仅0.2 μm,参与疼痛感觉传递的C纤维直径在0.1~0.2 μm之间;锥体细胞轴突的无髓鞘侧枝在局部皮层形成大量连接,其平均直径也仅0.3 μm。类似地,多数(棘或终扣型)中枢神经系统突触仅具有亚微米尺度。在这样小的长度尺度上,参与的分子数目也很少,因此噪声的影响就戏剧性地增加。我们这里所讨论的细胞水平噪声,主要是来源于神经系统的。我们讨论其神经元功能后果[2]。
在有噪声的条件下,神经网络如何维持其稳定活性?神经网络可以通过几个途径来影响总体噪声的水平(图8-2)[2]。
图8-2 网络中噪声的积累
此图显示了3个简单的例子。在这些例子中,具有等级性电位的神经元线性地总和了输入。(a)显示信号汇聚到单个神经元。如果进来的信号含有独立噪声,那么突触后神经元的噪声水平将按比例地取决于信号数目N的平方根(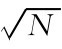 ),而测得的信号量与N成比例。如果信号中的噪声是完全相关的,那么神经元里的噪声也与N成比例。(b)显示信号经过一系列的神经元。在此情况下,噪声水平按相继的神经元数目之平方根的比例增加。与此相对比,通过神经网络的相互作用,平行连接(未显示在图上)并不增加噪声。(c)显示神经网络中的返回导致了相关联噪声的增加。(图引自[2])
),而测得的信号量与N成比例。如果信号中的噪声是完全相关的,那么神经元里的噪声也与N成比例。(b)显示信号经过一系列的神经元。在此情况下,噪声水平按相继的神经元数目之平方根的比例增加。与此相对比,通过神经网络的相互作用,平行连接(未显示在图上)并不增加噪声。(c)显示神经网络中的返回导致了相关联噪声的增加。(图引自[2])
各个神经元的其他计算操作可以改变网络噪声的积累。放大的线性操控使得信噪比不发生改变。非线性的操控如阈值的倍增,可以对噪声产生不同程度的影响。一般来说,倍增操控会增加输出的变异系数(CV),而阈值化则降低CV。有几个研究考察了噪声如何在神经元样的非线性系统里起作用。中枢神经系统高度平行、分布但又是致密的结构,可能帮助减少积累起来的噪声数量[2]。
实验性的证据提示,神经元活性的平均水平是由内稳态性可塑性机制维持的,这个机制动态地决定了突触强度、离子通道的表达或者神经递质的释放。这又转过来提示,神经元网络可以进行动态调节以削弱噪声的效应。此外,这些网络可能以某种方式连接,使个别神经元在反应特点方面大的变异,对网络行为仅有很小的效应[2]。
此外,在许多放电神经元中,增加一倍的输入导致少于两倍的输出。这提示,当信号通过神经元时,突触前噪声和细胞内噪声减弱。噪声维持得如此之小,这一事实提示,神经元网络的组构方式使得信号通过它们传播的时候,局部噪声的累积得以防止。这样,(以膜电位为基础)模拟性质的局部神经计算(神经元内的计算)以及(以动作电位为基础)更加数字化性质的总体宏观信息传输,可能是为建构具有噪声鲁棒性(noise-robust)的计算回路所需的基本要素[2]。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










