



孟德尔遗传学的进展
900年以后,遗传学的新成果出现
的速度在科学史上简直是史无前例的。查阅贝特森的教科书,我们就会为1900年后不久人们对孟德尔遗传学理解的成熟程度而感到惊讶。进步如此迅速的原因是什么?原因之一当然是这新学说本身的美满和简单足以吸引任何人去进行遗传试验来验证它是否普遍有效。由于这是一片新开拓的领域,几乎任何人都有取得新发现的机会。孟德尔定律可以对遗传方式作出预测并对这些预测立即加以检验。另一个原因则还没有定论,即认为1900年以前细胞学研究在35年中所取得的辉煌成就奠定了牢固基础,应当能够按细胞学,特别是从染色体的角度来说明几乎一切纯理论性的遗传学发现。染色体细胞学已成为通向生物学其他领域的桥梁,这座桥梁是在能够使用之前就已建成。然而奇怪的是,既使它在能使用之后也几乎被遗传学家,如在摩尔根之前的贝特森、凯塞尔、伊捷特等完全忽略了。
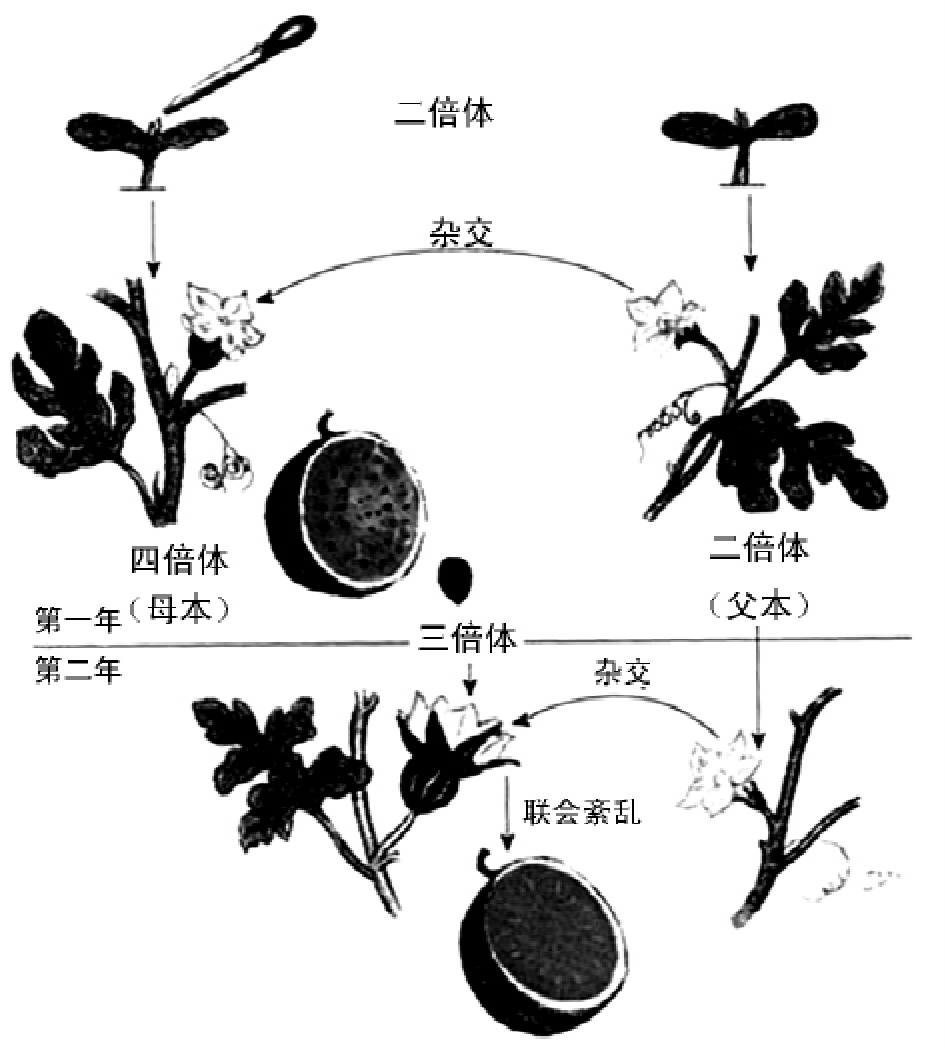
染色体的变化
遗传现象有关机制的知识被应用于生物学的各个不同领域,如进化生物学,发育生理学。下面将着重讨论传递遗传学方面。
一、半显性
在孟德尔所分析的七对性状中,他对每一对性状只分辨出两个变异体,即显性的和隐性的变异体。但是正如孟德尔本人所发现的那样,并不是所有的性状对都是如此。他曾经谈到花期就“几乎恰好处于亲本植株之间”。柯仑斯也同样发现某些因子并不是完全显性而是“半显性”,因而产生的F1表现型多少介于双亲之间。两年之后贝特森在用白家鸡和黑家鸡杂交时发现了蓝色的安道路西亚家鸡这样的半显性。
这不仅证实了半显性是存在的,而且还表明孟德尔定律对动物和植物都适用。大致在同一时期古诺根据对家鼠毛皮颜色基因的研究也论证了这一点。鉴于动物和植物的细胞及细胞核显示完全相同的现象这一事实,这一发现可能并非完全出乎意料。然而孟德尔的遗传定律同样适用于动物界和植物界的这一发现进一步摧毁了存在于动物学和植物学之间的古老界限。
二、基因——遗传单位
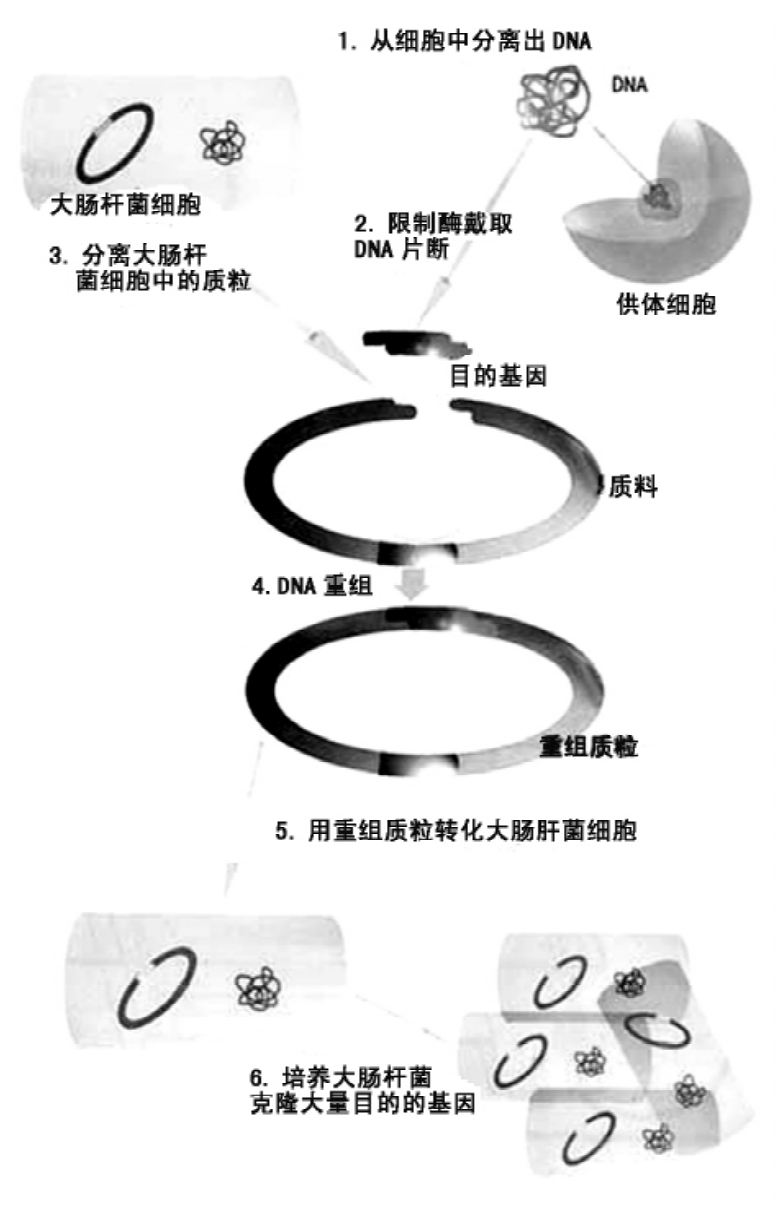
培养大肠杆菌的过程示意图
在1909年以前,对作为可见性状基础的遗传因子还没有一个公认的术语。斯宾塞、海克尔、达尔文、德弗里、魏斯曼以及其他考虑遗传现象的学者都假定存在着某种具有不同性质的颗粒物质,但是他们所使用的名称并未被广泛采用。
孟德尔在他的研究工作中对遗传物质本质的推论严格限制在最低限度,鉴于1865年当时对细胞核和染色体的了解还极其有限,他的这种作法是非常明智的。他在试验中所指的特征和性状基本上限于表现型层次,虽然他所用的符号A、Aa、a被普遍认为是指遗传型的结构。他在论文的结论中曾经使用“因子”这词达10次之多,其中有几次和我们现在使用的“基因”这词的含义十分相似,但他对遗传物质并没有清晰的概念。不管孟德尔真正是怎样想的,就早期的孟德尔主义者看来,他所阐述的就是我们现在所说的孟德尔遗传。
虽然魏斯曼曾经暗示了种质与体质之间的区别,但是直到1900年还没有“表现型”和“遗传型”这两个学术名词。就德弗里看来遗传物质和躯体并没有实质上的差异,因为他所设想的泛子可以自由地从细胞核移往细胞质。他认为泛子就是单位性状或基本性状。他主张对每一个独立遗传的性状就有一个单独的遗传基础。德弗里有时也把遗传要素称作“因子”,贝特森和摩尔根学派起初也采用了这一名称。
和德弗里相仿,贝特森也分不清作为基础的遗传因子与作为外表的表现型性状之间的区别。他将“单位性状”看作是“配子结构中二者择一非此即彼的”。为了能够指明这样的二者择一情况,例如豌豆中的圆豆或皱豆,贝特森引用了“相对形质”。然而他又不能区分躯体性状和它在配子中的定子。由于种种原因在1910年之前几乎普遍地达成了一种默契,即在遗传因子与性状之间存在着1∶1的关系。因此当提到某个单位性状时究竟是指遗传基础还是其表现都无关紧要。正是由于这种自动的默契促使凯塞尔提出了他的“污染学说”。
随着1900年以后遗传学活动的迅速增多,就有必要为可以独立遗传的性状的物质基础制订一个术语。丹麦遗传学家约翰逊发现孟德尔的因子在作用上与德弗里所提出的泛子很相似,因而在1909年建议将泛子这个字简化为“gene”,表示遗传性状的物质基础。约翰逊是一个物理主义者,他最后所想的是要为基因这个词的定义涂上先成论色彩。他指责“将基因作为物质性的,在形态上表示结构特征的概念,这概念对遗传学的稳步发展非常有害必须立即加以提防。”因而他并没有为基因下定义而只是说“基因可以用来作为计量单位。我们无权按达尔文的微芽或魏斯曼的生源体或定子或者其他类似的推论性概念将基因定义为某种形态结构。我们也无权去设想每一个特殊基因相当于特定的表现型性状或发育中生物的特征。”
这个定义反映了那个时期贯穿于生物学领域中的意见分歧。物理主义者要按力学观点解释一切。胚胎学者出于后生论传统也同样不愿意接受颗粒性基因,因为这使他们想起了先成论。摩尔根起初不愿意承认基因,或者至少是颗粒性基因,也正是由于这样的考虑。最后,还有本质论的某些影响,本质论是反对将物种的本质加以分割的。实际上,从摩尔根开始,经过穆勒到沃森和克里克一直越来越接近基因的结构概念。约翰逊创用的“基因”这个术语很快就被普遍采用,因为它满足了对表示遗传单位的术语的迫切需要。然而由于缺乏严格的基因定义,这就成为后来引起某些争论的部分原因。产生思想混乱的另一个原因是,几乎一直到现在学者们对基因的含义是什么意见也不一致。例如,当谈到果蝇的白眼基因时,有的学者认为指的是白眼等位基因,另一些人则认为是指白眼突变发生的基因座,也就是所有白眼等位基因的基因座。
为不可见的,亚显微结构的遗传单位创造“基因”这字开始,直到充分了解它的本质,这段道路是漫长的,也是艰苦的。无数的遗传学家实际上是把他们的毕生科学事业奉献在这一探索上。我们行将见到,最后发现作为基因发挥作用的高分子确实具有约翰逊所拒绝承认的那种结构上的复杂性和特异性。怎样去发现基因的秘密?开始的确是一个非常折磨人的问题。摩尔根及其同事十分正确地决定从研究改变了的基因即“突变”着手,他们认为这可能是一个有希望的嵌入楔子。
三、新变异的起源
由于重新发现了孟德尔分离定律,遗传性变异的起源问题就变得很突出。等位基因的存在要求能作出说明。达尔文假定变异不断被补充,以便自然选择有充分的选择对象可以发挥作用。然而他说不出变异的来源。这时已到了解开达尔文的这个谜的时候,但是孟德尔主义者在研究这个问题时起初进展并不大。实际上他们必须克服重重障碍。
主要障碍是当时研究变异的大多数学者仍然认为有两类变异。例如达尔文发现有“很多细微差异可以称之为个体差异”,后来称为个体变异,连续变异,或波动变异。他对这类变异的重要性的认识是他的进化学说的支柱之一。然而达尔文又承认“有些变异……可能是突然发生的、或一步发生的”并列举出转叉狗和安康羊作为这种突变的例子。贝特森将这类突变称为不连续变异。并承认这两类变异存在着悠久的历史并和柏拉图的本质概念有密切关系。本质承受小幅度的偶然性变异,而任何重大的偏差只能通过新本质的突然发生。当时认为这两类变异的起因完全不同,在进化中的作用也十分不同。
四、个体变异或连续变异
如果承认有软式遗传就不难解释个体变异。内部条件或环境影响的任何变化都能影响个体的性状并使之发生改变。正如达尔文所作的说明,“在由于条件变化使躯体的结构发生改变的情况下,由于用或不用的部位分别增强活动或降低活动,或者其他原因,从躯体结构发生了改变的部位脱落下来的微芽本身也会发生变化,当它们充分繁衍后就会发展成为新的,改变了的结构。”相信软式遗传的其他学者也采取了类似的解释。旧性状将分级成新性状,而这些分级性状之间的差别很小,表现为连续变异。如果新的遗传性变异起因于某种未知过程,它将同样承受软式遗传并分级成原先已存在的变异。这样一来就承认了物种的本质具有产生连续个体变异的能力,在解释上也就不成问题。动、植物育种家普遍持有环境能影响遗传变异性的观点。
1883年当魏斯曼放弃了软式遗传后,情况就发生了根本变化。如果“生活条件”不能引起新的变异甚至不能提高变异性,那么个体变异的原因是什么?魏斯曼和德弗里对此都没有提出理由充分的学说,早期的孟德尔主义者当时正集中注意力于不连续变异问题上,很少或根本不重视个体变异。怎样去调合不连续的孟德尔因子和连续变异之间的矛盾是使他们伤透脑筋的事。
不仅仅是由于缺乏合适的资料阻碍了这个问题的解决,而且默默地接受了许多错误概念也是重要原因。这些错误概念除了两类变异而外,还包括软式遗传,融合遗传,模式思想以及对遗传型和表现型混淆不清。面临着上述困难和错误概念,在当时还不可能直接着手解决连续变异和新变异起源的遗传学问题。问题的解决实际是通过研究不连续变异这一迂回方式,虽然这种方式的前提是认为不连续变异和连续变异完全无关。

魏斯曼提出种质连续性理论
五、不连续变异
古代人就早已知道在一个群体中偶尔会有个别个体与众不同,即超出了种群变异的正常标准。这在野生动物、家畜和栽培植物、甚至在人类中都曾发现。超出种群正常变异的任何变异体都是不连续变异的例子。在民间文学中都曾绘声绘色地描述过白化体,六指人以及各种畸形。15世纪和16世纪早期,当自然界被赋予巨大的“生殖能力”、即产生新奇事物的能力时,各种怪物都活灵活现地出现在各种图书上,其中大多数是真正的畸形动物,其他的则是纯粹幻想的生物,例如狮首人身之类的嵌合体。
1590年,海德堡的药剂师斯普伦格在他的药圃中发现一株叶子形状完全不同的五月白屈菜。他将繁育后的种子寄往各处,过了一段时间后在欧洲各个主要的植物标本室都有它的标本,17世纪的大多数植物书籍中也有对它的描述。这新的变异体一般被看作是白屈菜属的一个新种。310年以后,月见草属的一颗异常植株启示了德弗里,使他提出了一个新的重要进化学说。
在栽培植物中明显的异常变异体比较常见,实际上许多有名的园艺变种都是由这些变异体产生的。最有名的异常变异体的例子是所谓的反常整齐花。1741年林奈的一个学生从瑞典乌普沙拉带回了一株植物样品送给他,这植物在外观,特殊气味,花、花萼、花粉与种子的特殊颜色上乍一看和普通的蛋黄草完全相同。然而普通蛋黄草具有和金鱼草相似的典型不对称花,而反常整齐花则是具有5个突起的辐射对称花。林奈得出的结论是“这种新植物由本身种子繁殖,因而是一个新物种,并不是一开始就有的。”
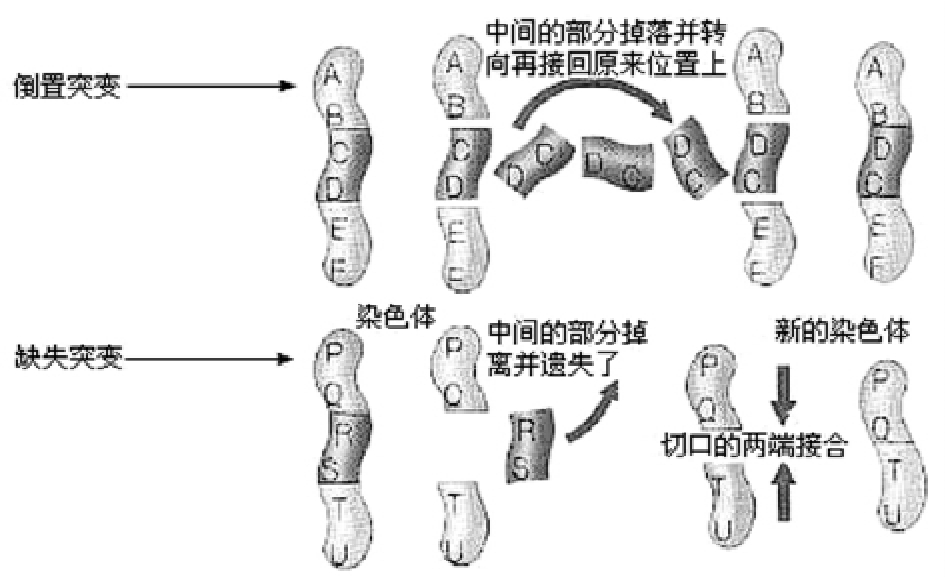
染色体变异
达尔文特别强调进化的渐进性,也就是说,连续变异在进化上的重要性,但这并没有使他的所有同时代人信服。赫胥黎,高尔敦,克里尔克及其他人,偏重通过不连续变异的骤变式新物种和模式起源。然而再也没有别的人比贝特森更加清楚地认识不连续变异的重要意义,他曾经收集了大量材料来证明他的论点。
六、德弗里和突变
1886年,德弗里在拉马克月见草的大种群中发现了两个植株,他认为这两个植株与所有其他个体极不相同可以看作是新产生的物种。它们在德弗里的试验园中经过自花授粉仍然极端稳定。从马铃薯地里移植到试验园的拉马克月见草的个体中也还有其他的新模式产生。后来除了许多次要的变异体而外,德弗里还发现了20株以上的个体可以认为是新种,在自花授粉后确实稳定不变。
德弗里为这类新“物种”产生的过程引用了“突变”这个词。考虑到这词在遗传学说中十分重要因而不妨多说几句。“突变”这个词早在17世纪中叶就被用来表示形体的剧烈变化。从一开始它就既用于不连续变异又用于化石的变化。1867年这词被瓦根正式引用于古生物学,指种系系列中可以分辨的最小变化而言。德弗里很了解这种用法因为他曾特地提到过瓦根。就像我们的语言中很多词一样,“突变”这词既用于过程又用于过程的结果。但是比这更复杂更容易混淆的是“突变”。有时用来指遗传型的变化,有时却指表现型变化。更糟的是,在德弗里心目中,突变是一种进化现象而在以后的遗传学史中它越来越成为专门的遗传现象。关于突变概念的这种混乱情况必须有所了解才能懂得为什么突变在进化中的作用一直长期争论不休。
虽然德弗里用“突变”这个词表达新种的突然出现,但是他并不了解这类变化的物理本质,而且事实上,他是将之用来表示表现型的突然变化。这已经被后来研究月见草的学者们证实,他们证明了德弗里所说的突变几乎全都是染色体重排的表现,其中很少是现代所指的基因突变。
经过几十年的遗传学研究才使“突变”这词摆脱了它原来的含义,正如德弗里所断言的,它是产生新种的过程的羁绊。虽然德弗里对他的突变所作的进化意义上的解释是错误的,但比起在他以前的任何人,他更强调新遗传性状的真正来源,在这一点上理应归功于他。后来,孟德尔以及研究遗传现象的其他学者就一直探索原已存在的遗传因子和性状的传递。德弗里促使人们注意遗传性新事物的来源问题。不管“突变”这个词的含义从1901年以来发生了多么大的变化,从那时起突变一直是遗传学的一个重要问题。
德弗里叙述了他是多么勤奋刻苦地寻求一种理想的植物来明确论证通过突变的物种骤然形成。他研究了一百多种植物,但是除了一种以外他将其余的全都放弃,因为它们的变异都不能像他所预期的那样保持下去。他曾强调指出月见草是多么特殊,然而他显然从来没有意识到,将一个新学说奠基于从单个特殊物种所观察到的现象上是多么危险。
正如瑞纳,克莱兰行,爱迪生以及其他遗传学者的出色研究所论证的那样,月见草有一套特殊的易位染色体系统。这系统由于纯合子的致死现象因而在杂合性上是永远平衡的。德弗里看作是突变的现象实际是这类染色体环的分离产物。这种情况在其他植物物种和动物中并不存在。德弗里的突变既不是正常变异的来源也不是物种形成的正常过程。然而他的“突变”这一术语却在遗传学中保留了下来,这是因为摩尔根保留了它,尽管摩尔根是将之转用于十分不同的遗传现象。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。






![评《哲学小辞典》中的[孟轲]](https://file.guayunfan.com/2020/zb_users/upload/2020/09/10.jpg)




