



第三节 HLA分子的分布、结构和功能
一、HLA分子的分布
HLA-Ⅰ类分子广泛分布于体内有核细胞表面,包括血小板和网织红细胞表面。成熟红细胞一般不表达Ⅰ类分子。淋巴细胞表面Ⅰ类分子密度最大,其次为肾、肝及心,密度最低的是肌肉和神经组织。
HLA-Ⅱ类分子主要分布于抗原递呈细胞(如B细胞、单核巨噬细胞、树突状细胞等)表面,激活的T细胞、内皮细胞也可表达Ⅱ类分子。
HLA-Ⅲ类分子主要是一些存在于血清及体液中的可溶性分子。包括补体成分(如C2、C4、B因子)、肿瘤坏死因子(TNF)、热休克蛋白(HSP)等。
二、HLA分子的结构
(一)HLA-Ⅰ类分子的结构
Ⅰ类分子是由2条多肽链组成的异二聚体,一条为由HLA-Ⅰ类基因编码的α链或称重链(44ku);另一条为非HLA基因编码的β2微球蛋白(β2m)或称轻链(12ku)。Ⅰ类分子根据其功能及结构可分为4个区(图5-2)。
1.肽结合区 由α链氨基端的两个约90个氨基酸残基的功能区α1和α2组成,是与抗原肽结合的部位,约容纳8~13个氨基酸残基。该区同时也是T细胞抗原受体识别的部位。
2.Ig样区 该区由α链的α3功能区和β2微球蛋白组成。α3区氨基酸顺序高度保守,与Ig的恒定区具有同源性。它是T细胞CD8分子的识别部位。β2微球蛋白不插入细胞膜而游离于细胞外,它与α2、α3功能区相互作用对维持Ⅰ类分子天然构型的稳定性有重要意义。
3.跨膜区 以螺旋形式穿过胞膜的脂质双层,将Ⅰ类分子锚定在膜上。
4.胞浆区 该区约30个氨基酸残基位于胞浆中,与细胞内外信息传递有关。
(二)HLA-Ⅱ类分子的结构
Ⅱ类分子是由两条结构相似的多肽链α链(34ku)和β链(29ku)组成的异二聚体,在细胞膜外分别形成α1、α2与β1、β2功能区。Ⅱ类分子也可分4个区(图5-2)。
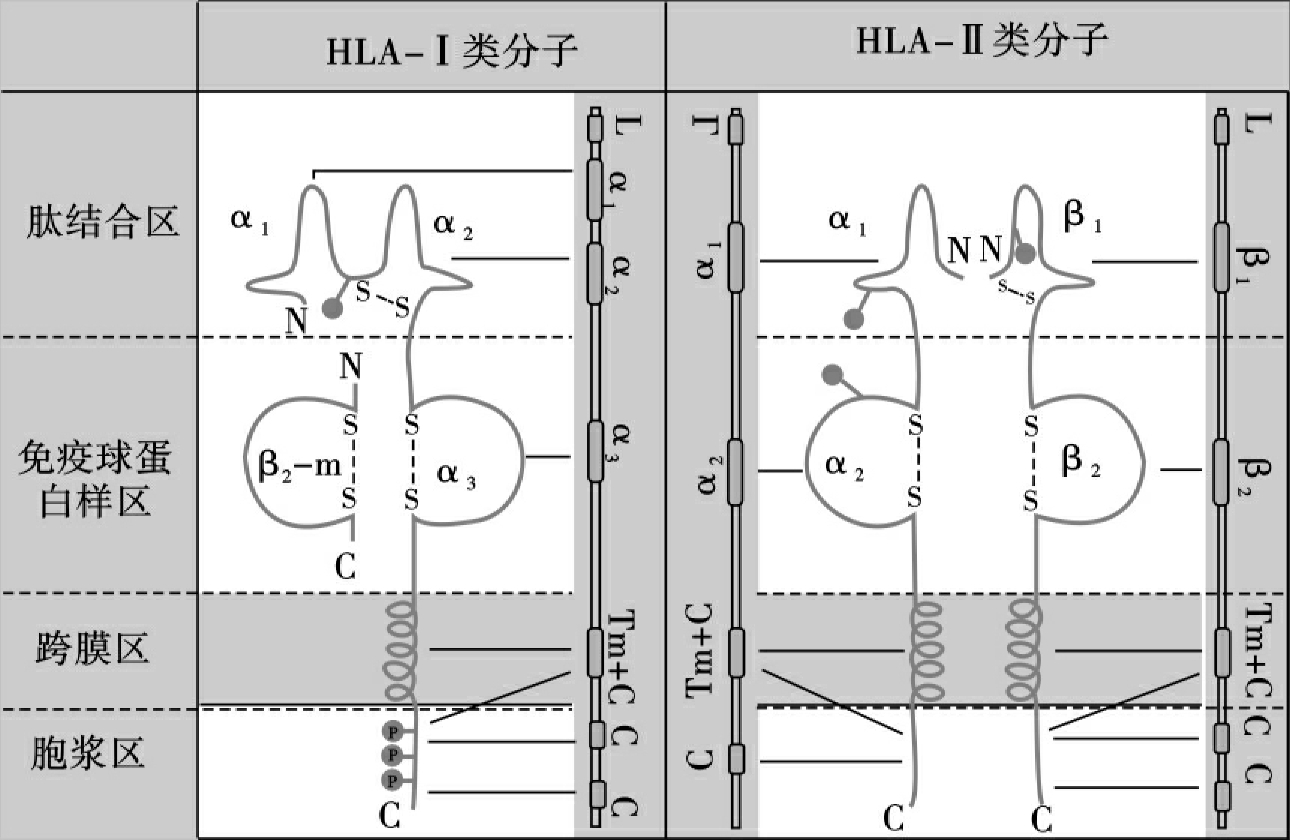
图5-2 HLA-Ⅰ类分子、HLA-Ⅱ类分子结构示意图
1.肽结合区 由α1和β1功能区组成,是抗原肽结合区,该区约容纳12~24个氨基酸残基。该区也是T细胞抗原受体识别部位。
2.Ig样区 由α2和β2功能区组成,此区是T细胞CD4分子的识别部位。
3.跨膜区 该区含有25个氨基酸残基,靠疏水结构穿过细胞膜的脂质双层,将Ⅱ类分子锚定在膜上。
4.胞浆区 含有10~15个氨基酸残基,游离于胞浆中,参与跨膜信号的传递。
三、HLA分子的功能
(一)参与抗原的识别和递呈
HLA分子的最主要功能之一是递呈抗原,参与免疫应答。两类HLA分子分别负责递呈不同类型抗原。外源性抗原与抗原递呈细胞内HLA-Ⅱ类分子结合,形成抗原肽-HLA-Ⅱ类分子复合物,递呈给CD4+T细胞;内源性抗原与靶细胞内HLA-Ⅰ类分子结合,形成抗原肽-HLA-Ⅰ类分子复合物,递呈给CD8+T细胞。
(二)引起移植排斥反应
HLA抗原呈现很强的免疫原性,在同种组织器官移植时,可在受者体内诱导产生相应的抗体和特异的T细胞从而攻击移植物细胞而发生排斥反应。
(三)参与T细胞限制性识别
在T细胞与抗原递呈细胞之间以及T细胞与靶细胞之间相互作用时,都只能识别自身HLA分子递呈的抗原肽而不能识别非己HLA分子所结合的相同的抗原肽。而且,CD8+T细胞必须同时识别内源性抗原肽和HLA-Ⅰ类分子才能被活化,称为HLA-Ⅰ类限制性;CD4+T细胞必须同时识别外源性抗原肽和HLA-Ⅱ类分子才能被活化,称为HLA-Ⅱ类限制性。
(四)参与T细胞分化过程
T细胞在胸腺内的发育成熟过程中,经历了阳性与阴性选择,使T细胞分化为具有自身耐受且受HLA限制的成熟T细胞。而HLA分子参与了阳性和阴性选择过程。
(五)参与免疫应答的遗传控制
机体对某种抗原能否发生免疫应答以及免疫应答的强弱是受遗传控制的,不同个体携带的HLA类型不同,对特定抗原的识别与递呈能力也不相同,从而实现机体对免疫应答的遗传控制。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。









